Revista de la Facultad de Ciencias
Agrarias. Universidad Nacional de Cuyo. Tomo 55(1). ISSN (en línea) 1853-8665.
Año 2023.
Original article
Growth and yield of Syagrus
schizophylla (Mart.) Glass. in response to light
gradients
Crecimiento
y rendimiento de Syagrus schizophylla (Mart.) Glass. en
respuesta a gradientes de luz
Janie Mendes Jasmim1
Rodollpho Artur de Souza
Lima1
Weverton Pereira Rodrigues1
Henrique Duarte Vieira1
Eliemar Campostrini1
1
Universidade Estadual do Norte Fluminense Darcy Ribeiro (UNEF). Laboratório de
Fitotecnia do Centro de Ciências e Tecnologias Agropecuárias (CCTA). Av.
Alberto Lamego. 2000. Parque Califórnia. Campos dos Goytacazes. Rio de Janeiro.
Brasil.
Abstract
This research studied
growth and yield of Syagrus schizophylla, an extinction-endangered
ornamental palm, grown under five light gradients. The treatments were: G1
- PAR=1234.10 μmol photons m-2 s-1,
G2
- PAR=913.16 μmol photons m-2 s-1,
G3
- PAR=666.34 μmol photons m-2 s-1,
G4
- PAR=419.56 μmol photons m-2 s-1 and G5 - PAR=534.77 μmol photons m-2s-1.
Before the experiment and at three, five and seven months of treatment, growth
(plant height, collar diameter, number of leaves, petiole length, leaf length
and width), gas exchange, chlorophyll a, and leaf green color intensity
were assessed. The highest net photosynthetic rates were observed in plants
under G2, G3
and G4.
Values of maximum quantum efficiency (Fv/Fm)
over 0.75 were observed under G2.
At seven months, estimated SPAD values were 36 in G2
plants and 32 in G1 plants. According to the Dickson quality index (DQI),
presented the highest growth and development rates.
We conclude that G2 suits plants to be
transplanted into the field, whereas G3 would be best for plants grown
under shade environments.
Keywords: Arecaceae; Gas Exchange; Palm tree; Photosynthesis; Shading.
Resumen
Syagrus schizophylla es una especie de palmera
ornamental en peligro de extinción. El objetivo de esta investigación fue
estudiar los aspectos fisiológicos del crecimiento y la calidad de las plantas
producidas bajo gradientes de luz. Los tratamientos de gradientes de luz
fueron: G1 - PAR = 1234.10 μmol fotones m-2 s-1,
G2 - PAR = 913.16 μmol fotones m-2 s-1,
G3 - PAR = 666.34 μmol fotones m-2 s-1),
G4 - PAR = 419,56 μmol fotones m-2 s-1 y G5 - PAR = 534,77 μmol
fotones m-2
s-1.
Antes de aplicar los tratamientos y a los tres, cinco y siete meses de
tratamiento, se analizaron la altura de la planta, diámetro del collar, número
de hojas, largo del pecíolo, largo y ancho de la hoja), intercambio gaseso,
fluorescencia de la clorofila a y color verde de la hoja. Las tasas
fotosintéticas netas más altas se observaron en plantas bajo G2, G3 y G4. Em G2
se observaron valores de Fv/Fm superiores a 0,75. En el séptimo mes, los
valores estimados del índice SPAD fueron 36 en plantas bajo G2 y 32 en plantas
bajo G1. El mejor crecimiento y desarrollo de las plantas se observó en G2 de
acuerdo con DQI que muestra que este gradiente debe usarse para plantas
destinadas al campo, mientras que G3 sería mejor para plantas destinadas a
entornos sombríos.
Palabras clave: Arecaceae; Intercambio gasesoso; Palmera; Fotosíntesis; Sombreado.
Originales: Recepción: 08/07/2022
Aceptación: 01/03/2023
Introduction
Syagrus schizophylla (Mart.) Glass. is an indigenous Arecaceae, naturally found in the Brazilian
Sandy Coast (Restinga) and the Atlantic Rain Forest (17).
Ordinarily known as aricuriroba, coco-babão and licuriroba, the
fruits are considered an unconventional food and an alternative source of carbohydrates,
proteins and lipids (23). Unfortunately,
the species is considered endangered by the Flora Conservancy National Center
(CNCFlora Red List, Brazil), mainly given to both urban and tourist pressures (25).
In forests, palm
distribution depends on light and water availability. The Arecaceae family
shows a wide diversity of genera and species, still unstudied regarding
adaptive traits. Due to spatial distribution and diversity, palms are suitable
models for studying biodiversity of tropical and subtropical ecosystems (17, 23, 25).
The photosynthetically
active radiation (PAR, μmol fótons m-2s-1), ranges between 400
and 700 nm (10). Red light incidence
(600-700 nm) on the leaf mesophyll enhances stomatal opening (1). However, exposure for long periods may cause
red light syndrome, leading to low quantum yield, low photosynthetic capability
and lack of stomatal response (20, 27).
Light intensity and quality vary according to daytime, season, location,
climate, position within the plant canopy, and even within the cell (21). Such variations can result in
morphoanatomical changes affecting leaf biomass allocation, chlorophyll a/b
ratio, leaf thickness, stomatal density and photochemical dissipation (27), organization of the photosynthetic
apparatus, etiolation, leaf area and carbohydrate storage, among others (14, 20). Blue, red and infrared light activate
specific receptors, triggering independent key physiological events (20). This research hypothesizes that using shade
nets with light gradients in the blue and red ranges would result in optimized
plant carbon allocation and photosynthesis. We studied the physiological
response of S. schizophilla to different wavelengths determining the
best environmental conditions for sapling yield. The experiments used different
shade nets, assuming that specific wavelengths modify the intercepted energy,
leading to optimized carbon fixation, growth and yield (5, 10).
Given the need for the preservation and recovery of negatively
anthropized areas, this study might contribute to optimizing plant
conservation, preservation and sustainable use, while offering income
diversification in rural areas.
Material
and methods
Plant
material, seedling transplant and light gradients
Seeds were obtained and
germinated as described by Beltrame et al. (2019).
Ninety days after emergence, seedlings of S. schizophylla were
transplanted into pots of approximately 6.3 L, previously filled with a mixture
of commercial substrate Plantmax® and coconut fibre (1:1; v:v) with the
following physicochemical characteristics: P = 0.01 g kg-1,
K = 5.47 g kg-1,
S = 4.82 g kg-1,
Ca = 7.89 g kg-1,
Mg = 4.34 g kg-1;
B = 15.07 mg kg-1,
Fe = 1.54 mg kg-1,
Mn = 4.34 mg kg-1,
Zn = 26.46 mg kg-1,
Cu = 0.2 μg kg-1,
pH = 4.11, CE 0.8 μS cm-1,
bulk density = 0.42 g cm-3,
true density = 1.58 g cm-3 and total porosity of 73% of
container volume.
Initially, the plants were
kept for 270 days under an average of 534.77 μmol photons m-2 s-1 (May 2016 to February 2017),
in a greenhouse, with mean PAR 534.77 μmol photons m-2 s-1.
After that period, treatments lasted 210 days (from March to October 2017). The
seedlings were randomly organized and grown under tunnels (1.80x1.50x1.80 m) covered
with different shade nets resulting in different light gradient treatments:
plain sunlight - control (G1)
(PAR=1234.10 μmol photons m-2 s-1),
red Chromatinet® 50% of shade (G2)
(PAR=913.16 μmol photons m-2 s-1),
two overlapping layers of red Chromatinet® 50% of shade (G3)
(PAR=666.34 μmol photons m-2 s-1),
black polyolefin 50% of shade (G4)
(PAR = 419.56 μmol photons m-2 s-1)
and overlapping layers of milky plastic film and polyolefin 50% of shade (G5) (PAR = 534.77 μmol
photons m-2
s-1).
HOBO Pro v2 Data Loggers
hourly monitored mean, minimum, and maximum temperatures throughout the
experimental period. In addition, light spectral quality was evaluated using
the USB2000+RAD Ocean Optics UV/Vis spectrum radiometer, obtaining three
consecutive readings in each tunnel, at 9 am and three consecutive readings at
noon, on bright sunny days.
Growth analyses
Before light treatments
(BT), and at three, five and seven months, we measured collar diameter (DC),
shoot height (SH), number of leaves (NL), petiole length (LP), leaf length (LL)
and width (LW). The LP, LL and LW were measured with a ruler, on the second
pair of fully expanded leaves.
After seven months, shoot
(SDW) and root dry weight (RDW), and plant total leaf area (TLA) were
determined. All leaves were detached, and leaf blades and petioles were
separated with pruning shears for TLA determination using a Li-3100 (Li-Cor,
USA) leaf area meter. For dry weight determination, plant shoot and roots were
separated, paper bagged and dried in a convection oven at 70 ± 2°C for 96
hours. Root and shoot dry weights were gravimetrically determined (±0.0001 g).
Dickson quality index
(DQI) was calculated according to Dickson et al. (1960) (eq. 1), using total dry mass (TDM),
shoot height (SH), collar diameter (DC), shoot dry weight (SDW) and root dry
weight (RDW).
![]()
Gas exchange, chlorophyll a fluorescence and green color
intensity
Gas exchange was evaluated
between 8 am and 10 am with a portable infrared analyzer (IRGA - model Li-6400
XT - Li-Corporation/USA). Evaluation cycles correspond to cycle 1 = one month after
initializing treatments (AT)], cycle 2 = three months after AT and cycle 3 =
seven months after AT. Net photosynthesis (A), transpiration (E),
stomatal conductance (gs)
and internal vs. external CO2 (Ci/Ca), were determined
on the second pair of completely developed leaves. For that purpose, a 6 cm2
chamber was conditioned with 1500 μmol photons m-2 s-1 light intensity, 500 μmol s-1
airflow, and 400 ppm standard CO2 concentration (obtained with a
CO2
mixer) at room temperature, with mean temperatures varying
between 25 and 30°C. Light response curves with 24 levels of PPFD from 1500 to
0 μmol m-2
s-1 allowed for Optimal
photosynthetic photon flux density (PPFD) determination. Meanwhile, and on the
same leaf used for gas exchange measurements, chlorophyll fluorescence was
determined using a Pocket fluorimeter PEA (Plant Efficiency Analyser,
Hansatech, England).
Leaves were dark-adapted
for 30 minutes with leaf clips (Hansatech), avoiding leaf veins, so that all
reaction centres were in the oxidized state. Maximum quantum yield of
photosystem II (Fv/Fm) and the photosynthetic index (PI) were
determined according to Strasser et al. (2004).
Leaf green colour intensity (SPAD index), which correlates with chlorophyll
content, was measured with a portable chlorophyll meter (model SPAD-502
Minolta, Japan). Mean values were obtained from eight measures per plant.
Gas exchange and
chlorophyll fluorescence were measured one, three and seven months later. SPAD
measurements were taken before treatments (AT) and three, five and seven months
later.
Statistical analysis
The experiment was
conducted in a completely randomized design with five light gradient treatments
and six replications, totalizing 30 plants. Data were subjected to ANOVA, and
means were compared by Tukey test at 5% probability. Statistical analyses were
performed with R (26).
Results and discussion
Light, gas exchange and photosynthetic capacity
Under all light gradients, air
temperature and humidity were very similar, with mean temperatures varying
between 25 and 30°C, and air humidity around 80%. Light spectrum varied as
follows: 300 - 900 nm in G1,
300 - 850 nm in G2,
300 - 800 nm in G3,
400 - 700 nm in G4 and 400 - 750 nm in G5. Net photosynthesis (A)
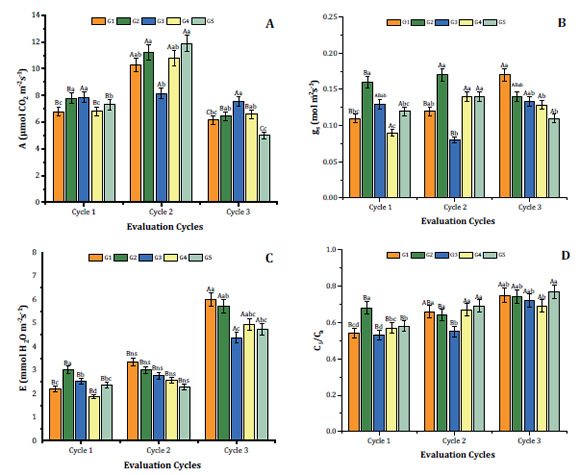
varied independently of light gradient (Figure 1A).
Cycle 1 (one month of light treatment), cycle 2 (three months of
light treatment) and cycle (seven months of light treatment). Upper case
letters compare evaluation cycles; lower case letters compare light gradients,
by Tukey test (p<0.05).
Ciclo 1 (un mes de fototratamiento), ciclo 2 (tres meses de
fototratamiento) y ciclo (siete meses de fototratamiento). Letras mayúsculas
comparan ciclos de evaluación; letras minúsculas comparan gradientes de luz, según
la prueba de Tukey (p<0,05).
Figure 1: Net photosynthesis (A), stomatal conductance (gs), transpiration (E)
and internal vs. external concentration of CO2
(Ci/Ca) of Syagrus
schizophylla plants under light gradients (G1,
G2, G3, G4 and G5)
and evaluation cycles.
Figura 1: Fotosíntesis neta (A), conductancia estomática (gs),
transpiración (E) y concentración interna vs. externa
de CO2
(Ci/Ca) de plantas de Syagrus schizophylla bajo gradientes
de luz (G1, G2, G3, G4 and G5) y ciclos de evaluación.
At cycle 1, the highest A values
were observed in G2 and G3, reaching approximately 8.0
μmol CO2
m-2 s-1 (Figure
1A).
At cycle 2, that is, after
three months of light treatment, mean A values were different among light
gradients. G2 and G5 were different from G3, showing the highest rates of net
photosynthesis (Figure 1A). At seven months (cycle 3),
higher mean values were observed in plants from G2, G3 and G4, varying between
6 and 7.5 μmol CO2 m-2 s-1 (Figure 1A). Gas exchange measurements, particularly net
photosynthesis, allows understanding genotype x environment photosynthetic
patterns (15), constituting a reliable
indicator of plant physiological status (24).
Stomatal conductance (gs) showed statistical
differences among treatments. At evaluation cycle 1, the highest mean gs were observed in plants
from G2
and G3 (Figure 1B). At cycle 2, no significant differences were observed in
G2, G4
and G5.
The lowest gs was
observed in G3 (Figure 1B). However, at evaluation cycle 3, mean gs exceeded 0.11 mol m-2
s-1 regardless of light gradient (Figure 1B).
While studying S.
schizophylla palm plants, the highest gs observed in G2, G3 (cycles 1
and 3) and G2 (cycle 2), suggested that red light somehow influences the
stomatal opening. Our results might be related to light incidence and quality,
resulting in fast gs increases
and responses to white light components (1).
Dumont
et al. (2013) reported that stomata are
especially sensitive to blue light. In this sense, a weak but significant
linear correlation between A and gs (R = 0.45), corroborated
our results.On the other hand, Lavinsky et al.
(2014) reported that A increased 3.5 fold in Euterpe
edulis saplings acclimated at 25.0 mols photons m-2 d-1,
in relation to understory saplings, acclimated at 1.3 mol photons m-2
d- 1,
and presenting low respiration rate and rapid gas response.
Transpiration rate (E)
of S. schizophylla plants showed statistical differences among light
gradients at evaluation cycles 1 and 3. At evaluation cycle 1, E was highest in
G2
(Figure 1C). No significant differences
in E under light gradients were observed in cycle 2. At cycle 3, the highest E
values were close to 6.0 mmol H2O
m-2
s-1 in G1 and G2 plants (Figure
1C). Assimilation of CO2 inevitably requires water loss
(E), as gas diffusion rates increase with stomatal conductance (18).
Mean internal vs. external CO2 ratio (Ci/Ca) of S.
schizophylla plants showed statistical differences, with the highest Ci/Ca in G2 plants (Figure 1D). At evaluation cycle 2, mean Ci/Ca
ratios in G1,
G2, G4
and G5 showed no differences (Figure 1D). In cycles 2 and 3, at
three and seven months, mean Ci/Ca
ratio increased significantly under light treatments,
probably related to plant growth and development, and light quality, since gas
exchange is maximized under blue (400-500 nm) and red (600-700 nm) lights (25). In this study, S. schizophylla plants
showed different responses under different shading, stating a species-dependent
need to use shade nets.
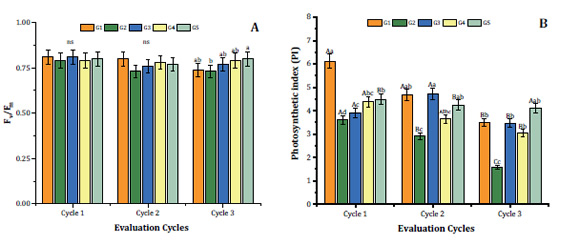
Cycles 1 and 2 showed no
statistical differences among Fv/Fm
(Figure 2A).
Upper case letters compare evaluation cycles; lower case letters
compare gradients by Tukey test (p < 0.05). *ns = non-significant (p <
0.05).
Las letras mayúsculas comparan los ciclos de evaluación; las
letras minúsculas comparan gradientes mediante la prueba de Tukey (p <
0,05). * ns = no significativo (p < 0,05).
Figure 2: Maximum quantum yield of
photosystem II (Fv/Fm) and photosynthetic
index (PI) of Syagrus schizophylla plants under light gradients
(G1, G2, G3,
G4
e G5)
at evaluation cycles 1 (one month of light treatment), 2 (three months of light
treatment) and 3 (seven months of light treatment).
Figura 2: Rendimiento cuántico máximo del fotosistema II (Fv/Fm) e índice
fotosintético (PI) de plantas de Syagrus schizophylla bajo gradientes
lumínicos (G1, G2, G3, G4 y G5) en los ciclos de evaluación 1 (a un mes de
tratamiento con luz), 2 (a los tres meses de tratamiento con luz) y 3 (a los
siete meses de tratamiento con luz).
In contrast, cycle 3
resulted in Fv/Fm
significant differences among G2 (lower value) and G5
(higher value) plants, all exceeding 0.75 (Figure 2A). These results suggest that despite the light treatment, S.
schizophylla did not suffer photoinhibition, in accordance with Bolhàr-Nordenkampf et al. (1989) who reported that Fv/Fm
varied from 0.75 to 0.85 in plants with intact photosynthetic
apparatus. These authors state that photoinhibitory damage on PSII reaction
centres causes decreased Fv/Fm. Lavinsky et al. (2014)
reported that E. edulis acclimated to understory conditions showed a
steep drop in Fv/Fm, from 0.8 to 0.5,
during the first week after being transferred to a more luminous environment.
After 21 days of re-acclimation, Fv/Fm increased, and at 110
days it was almost re-established to the initial value (0.7). Another study
also observed that stressed plants tend to decrease Fv/Fm
values, dissipating the exceeding energy and preventing
photoinhibition (12). In accordance with
our findings, other authors found decreased Fv/Fm
and photosynthetic index (PI) in cucumber plants
exposed to red light (19, 20, 27). Osório et al. (2012)
also state low Fv/Fm for photosynthesis,
photochemical efficiency and photoinhibition studies.
Regarding photochemical
efficiency, the photosynthetic index (PI) showed interaction among
treatments. The highest PI value at cycle 1 exceeding 6, was observed in
plants from G1,
while other treatments had PI values under 4.5 (Figure
2B). At evaluation cycle 2, the highest values
were observed in G1,
G3
and G5 (Figure 2B). Cycle 3, showed no significant differences among PI in
G1, G3, G4 and G5,
varying from 3.5 to 4.5 (Figure 2B). Finally, excepting G5 plants, PI of S.
schizophylla plants decreased along cycles, probably given by light stress (13).
Plant growth and development as a function of light
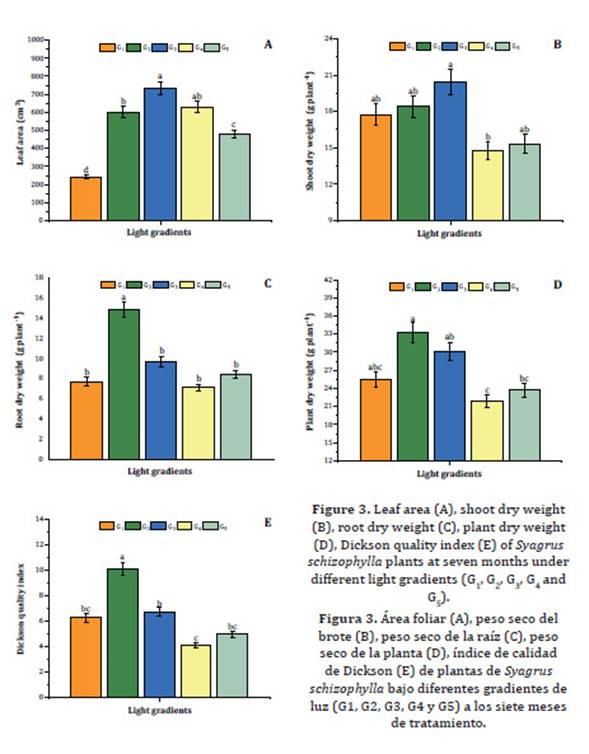
Total leaf area was higher
in G2, G3
and G4,
followed by G5 and G1 plants; the latter presenting
the smallest leaf area, 250 cm2 (Figure 3A).
Different letters indicate significant differences for Tukey test
(p < 0.05). *ns = non-significant (p < 0.05).
Diferentes letras indican diferencias significativas según la
prueba de Tukey (p < 0,05). * ns = no significativo
(p < 0,05).
Figure 3: Leaf area (A), shoot dry
weight (B), root dry weight (C), plant dry weight (D), Dickson quality index
(E) of Syagrus schizophylla plants at seven months under different light
gradients (G1,
G2, G3, G4 and G5).
Figura 3: Área foliar (A), peso seco del brote (B), peso seco de la raíz
(C), peso seco de la planta (D), índice de calidad de Dickson (E) de plantas de
Syagrus schizophylla bajo diferentes gradientes de luz (G1, G2, G3, G4 y
G5) a los siete meses de tratamiento.
Similarly, Gatti et al.
(2011) observed decreasing leaf area of E.
edulis plants as light intensity increased.
Significant differences
were only observed among shoot dry weights in G3 and G4 (Figure 3B). However, the highest root dry weight, 14.5 g plant-1, was observed in G2
(Figure 3C).
Mean plant dry weight under G2 exceeded 33 g plant-1
(the highest mean), while the lowest mean was 22.5 g plant-1, in G4 (Figure 3D).
The highest Dickson
quality index (DQI) was observed under G2 (above 10.0); followed by G3, G1,
G5
and G4 plants, in decreasing order
and ranging from 7.0 to 4.0 (Figure 3E). DQI is considered a good
parameter for quality assessment in nursery plants, considering the amount and
even the distribution of phytomass, pondering different parameters that help
estimate plant performance in the field (9).
De Oliveira et al. (2009) observed DQI ranging from 2.29 to 2.77 for Copernicia
hospita seedlings after three months of growth, and values from 15.78 to
20.70 after nine months of growing in different-size containers under plain
sunlight. For B. capitata, Costa et al.
(2018) recommended red shade nets, inducing stomatal
and leaf anatomical changes enhancing light harvesting, photosynthesis and
plant growth and development. B. capitata plants grown under red shade
net (50%) showed higher biomass than plants from other shade treatments (4).
The present research suggests
that red shade nets contribute to increased plant growth rates and vigour of S.
schizophylla by enhancing photosynthetic efficiency. Moreover, gas
exchange, growth and development of S. schizophylla plants throughout
the experiment with light gradients generated valuable information in relation
to productivity. Plants exposed to a greater amount of light had greater
biomass. Additionally, in G2,
a higher DQI suggested this index may also predict plant survival after field
transplanting and/or shade environments. The results not only confirm that this
species is tolerant to light variations but also suggest it undergoes
acclimation, optimizing resource allocation through structural and
physiological adaptations.
Conclusions
One layer of Red Chromatinet®
50% should be recommended for the first year of growth of S.
schizophylla plants to be later transplanted to the field. Two Red
Chromatinet® 50% layers would be best for plants meant to grow under shade
environments.
Acknowledgements
To
the Coordination for the Improvement of Higher Education Personnel (CAPES) for
granting a scholarship to the first author.
1.
Aasamaa, K.; Aphalo, P. H. 2015. Effect of vegetational shade and its
components on stomatal responses to red, blue and green light in two deciduous
tree species with different shade tolerance. Environmental and Experimental
Botany. 121: 94-101. 10.1016/j.envexpbot.2015.01.004.
2. Beltrame, R. A.; Jasmim, J. M.; Vieira, H. D. 2019.
Morphological characterization and germination of Syagrus schizophylla (Mart.)
Glass. (Arecaceae). Comunicata Scienciae. 10:54-64. https://
doi.org/10.14295/cs.v10i1.2997
3.
Bolhàr-Nordenkampf, H. R.; Long, S. P.; Baker, N. R.; Oquist, G.; Schereiber,
U.; Lechner, G. 1989. Chlorophyll fluorescence as probe of the photosynthetic
competence of leaves in the field: A review of current instrument. Functional
Ecology. 3: 497-514. https://doi. org/10.2307/2389624
4.
Costa, B. N. S.; Costa, I. J. S.; De Souza, G. A.; Dos Santos, D. N.; Da
Silveira, F. A.; De Melo, E. T.; Martins, A. D.; Pasqual, M.; Setotaw, T. A.;
Rodrigues, F. A. 2018. Anatomical modifications of Butia capitata propagated
under colored shade nets. Anais da Academia Brasileira de Ciências. 90:
3615-3624. https://doi.org/10.1590/0001-3765201820170347
5.
Costa, I. J. S.; Costa, B. N. S.; De Assis, F. A.; Martins, A. D.; Pio, L. A.
S.; Pasqual, M. 2018. Growth and physiology of jelly palm (Butia capitata)
grown under colored shade nets. Acta Scientiarum. Agronomy. 40:1-8.
https://doi.org/10.4025/actasciagron.v40i1.35332.
6.
De Oliveira, A. B.; Medeiros Filho, S.; Bezerra, A. M. E.; Bruno, R. L. A.
2009. Emergência de plântulas de Copernicia Hospita Martius em função do
tamanho da semente, do substrato e ambiente. Revista Brasileira de Sementes.
31: 281-287. https://doi.org/10.1590/S0101-31222009000100031
7.
Dickson, A.; Leaf, A. L.; Hosner, J. F. 1960. Quality appraisal of white spruce
and white pine seedling stock in nurseries. The Forestry Chronicle. 36: 10-13.
https://doi.org/10.5558/tfc36010-1
8.
Dumont, J.; Spicher, F.; Montpied, P.; Dizengremel, P. 2013. Effects of ozone
on stomatal responses to environmental parameters (blue light red light, CO2 and
vapour pressure deficit) in three Populus deltoides x Populus nigra.
Environmental Pollution. 173: 85-96. https://doi.
org/10.1016/j.envpol.2012.09.026
9.
Fonseca, E. P.; Valéri, S. V.; Miglioranza, E.; Fonseca, N. A. N.; Couto, L.
2002. Padrão de qualidade de mudas de Trema micrantha (L.) Blume,
produzidas sob diferentes períodos de sombreamento. Revista Árvore. 26:
515-523. https://doi.org/10.1590/S0100-67622002000400015
10.
Fujiwara, K.; Kozai, T. 1995. Physical microenvironment and its effects. In:
Aitken-Christie, J; Kozai, T; Smith, MAL (eds). Automation and environmental
control in plant tissue culture. Kluwer academic publishers. Netherlands. 50 p.
11.
Gatti, M. G.; Campanello, P. I.; Goldstein, G. 2011. Growth and leaf production
in the tropical palm Euterpe edulis: Light conditions versus developmental
constraints. Flora. 206: 742-748. https://doi.org/10.1016/j.flora.2011.04.004
12.
Gomes, F. P.; Oliva, M. A.; Mielke, M. S.; De Almeida, A. A. F.; Leite, H. G.;
Aquino, L. A. 2008. Photosynthetic limitations in leaves of Young Brazilian
Green Dwarf coconut (Cocos nucifera L. ‘nana’) palm under well-watered
conditions or recovering from drought stress. Environmental and Experimental Botany . 62: 195-204. https://locus.ufv.br//handle/123456789/23666
13.
Gonçalves, J. F. C.; Silva, C. E.; Guimarães, D. G.; Bernardes, R. S. 2010.
Análise dos transientes da fluorescência da clorofila a de plantas
jovens de Carapa guianenses e de Dipteryx odorata submetidas a
dois ambientes de luz. Acta Amazonica. 40: 89-98.
14.
Guidi, L.; Brunetti, C.; Ferrini, A.; Gori, A.; Tattini, M. 2016. UV radiation
promotes flavonoid biosynthesis, while negatively affecting the biosynthesis
and the de-epoxidation of xanthophylls: Consequence for photoprotection? Environmental and
Experimental Botany .
127: 14-25. https://doi.org/10.1016/j.envexpbot.2016.03.002
15.
Larcher, W. 2006. Ecofisiologia vegetal. São Carlos: Rima Artes e Textos. 531
p.
16.
Lavinsky, A. O.; Gomes, F. P.; Mielke, M. S.; França, S. 2014. Photosynthetic
acclimation in shade-developed leaves of Euterpe edulis Mart (Arecaceae)
after long-term exposure to high light. Photosynthetica. 52:351-357.
http://dx.doi.org/10.1007/s11099-014-0038-5
17.
Leitman, P.; Soares, K.; Henderson, A.; Noblick, L.; Martins, R. C. 2015. Syagrus
in Flora do Brasil 2020 em construção. Jardim Botânico do Rio de Janeiro. http://www.floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB15746 . Access in: 12 nov. 2016.
18.
Lu, Y.; Duursma, R. A.; Medlyn, B. E. 2016. Optimal stomatal behaviour under
stochastic rainfall. Journal of Theoretical Biology. 394: 160-171.
https://doi.org/10.1016/j.jtbi.2016.01.003
19.
Miao, Y.; Wang, X.; Gao, L.; Chen, Q.; Qu, M. 2016. Blue light is more
essential than red light for maintaining the activities of photosystem II and I
and photosynthetic electron transport capacity in cucumber leaves. Journal of
Integrative Agriculture. 15: 87-100. 10.1016/S2095-3119(15)61202-3
20.
Miao, Y.; Chen, Q.; Qu, M.; Gao, L.; Hou, L. 2019. Blue light alleviates ‘red
light syndrome’ by regulating chloroplast ultrastructure, photosynthetic traits
and nutrient accumulation in cucumber plants. Scientia Horticulturae
-Amsterdam. 257: 1-9. 10.1016/j.scienta.2019.108680
21.
Murchie, E. H.; Niyogi, K. K. 2011. Manipulation of photoprotection to improve
plant photosynthesis. Plant Physiology. 155:86-92.
https://doi.org/10.1104/pp.110.168831
22.
Osório, M. L.; Osório, J.; Gonçalves, S.; David, M. M.; Correia, M. J.; Romano,
A. 2012. Carob trees (Ceratonia siliqua L.) regenerated in vitro can
acclimatize successfully to match the field performance of seed-derived plants.
Trees. 26: 1837-1846. https://doi.org/10.1007/s00468-012-0753-0
23. Padilha, M. R. F.; Shinohara, N. K. S.; Macêdo, I. M. E.;
Bernardino, A. V. S.; Rocha, N. S.; Machado, J. 2017. Syagrus schizophylla: Unconventional
Food Plant of the Caatinga Biome with a high caloric value. Geama. 2: 53-57. http://www.journals.ufrpe.br/index.php/geama/article/view/1397
24.
Pires, T. P.; Souza, E. S.; Kuki, K. N.; Motoike, S. Y. 2013. Ecophysiological
traits of the macaw palm: A contribution towards the domestication of a novel
oil crop. Industrial Crops and Products. 44:200-210.
https://doi.org/10.1016/j.indcrop.2012.09.029
25.
Prieto, P. V.; Messina, T. 2012. Syagrus schizophylla in Lista Vermelha
da flora brasileira versão 2012. Centro Nacional de Conservação da
Flora. http://cncflora.jbrj.gov.br/portal/pt-br/profile/Syagrusschizophylla . Access in: 19 oct. 2016.
26.
R Core Team. 2015. A language and environment for statistical computing. R
Foundation for Statistical Computing. Vienna, Austria. http://www.R-project.org/ Access in: 22 jul.
2014.
27.
Rehman, M.; Ullah, S.; Bao, Y.; Wang, B.; Peng, D.; Lijun, L. 2017.
Light-emitting diodes: whether an efficient source of light for indoor plants?
Environmental science and pollution research. 24: 24743-24752.
10.1007/s11356-017-0333-3.
28.
Strasser, R. J.; Tsimilli-Michael, M.; Srivastava, A. 2004. Analysis of the
fluorescence transient. In: George C Papageorgiou; C Govindjee (Eds.).
Chlorophyll fluorescence: A signature of photosynthesis. Springer. 321-362.