Revista de la Facultad de Ciencias
Agrarias. Universidad Nacional de Cuyo. Tomo 56(1). ISSN (en línea) 1853-8665.
Año 2024.
Original article
Effects
of geomorphology and distribution of water sources for livestock on the
floristic composition and livestock receptivity of the Arid Chaco
Efecto
de la geomorfología y la distribución de las fuentes de agua para el ganado en
la composición florística y receptividad ganadera del Chaco Árido
1Consejo
Nacional de Investigacion Científica y Técnica (CONICET).
2Instituto
Nacional de Tecnología Agropecuaria (INTA). Estacion Experimental Agropecuaria
San Juan Calle 11 y Vidart 5427. Villa Aberastain. San Juan. Argentina.
3Universidad
Nacional de San Juan (UNSJ). Facultad de Ingenieria. Av. Lib. San Martín
(Oeste) 1109. C. P. A. J5400ARL.
4Universidad
Nacional de San Juan (UNSJ). Facultad de Ciencias Exactas, Físicas y Naturales
(FCEFyN). Cátedra de Manejo de Bosques y Pasturas Naturales. Gabinete de
Geología Ambiental. Av. Ignacio de la Roza 590 (O). Complejo Universitario
“Islas Malvinas”. Rivadavia. C. P. A. J5402DCS. San Juan. Argentina.
*joposcaglia@gmail.com
Abstract
Livestock
production in semi-arid areas is possible due to the presence of permanent water
sources, which create a radial pattern of grazing intensity known as the
piosphere. For this reason, we predicted that permanent water sources would
negatively impact the ecological conditions of plant communities, leading to
variations in livestock receptivity. To test this prediction, we compared
grazing gradients in two geomorphological units, using distance to water
sources as an indicator of accumulated livestock pressure. We assessed
variations in the botanical composition of both areas by analysis of variance
and principal components analysis. Additionally, we modeled the relationship
between distance to water source and livestock receptivity. Our results
revealed significant differences in the contribution of different species based
on their distance to water sources. Notably, a non-linear regression model
provided the best fit for the relationship between water source and livestock
receptivity in both geomorphological units. These findings demonstrate that the
distance to permanent water sources serves as a reliable indicator of
accumulated livestock pressure in semi-arid regions like the study area.
Keywords: arid Chaco,
natural, grasslands, piosphere, geomorphology
Resumen
La producción de
ganado en zonas semiáridas es posible debido a la existencia de fuentes de agua
permanentes, lo que genera un patrón radial de intensidad de pastoreo llamada
piosfera. Por tal motivo, nuestra predicción se basa en que las fuentes de agua
permanente influyen negativamente en las condiciones ecológicas de las
comunidades vegetales provocando diferentes receptividades ganaderas.
Comparamos gradientes de pastoreo tomando la distancia a las fuentes de agua
como indicador de presión ganadera acumulada en dos unidades geomorfológicas.
La variación en la composición botánica de las diferentes áreas se realizó
utilizando análisis de la varianza y un análisis de componentes principales. Se
efectuó un modelo de la relación entre la distancia a la fuente de agua y la
receptividad ganadera. Nuestros resultados mostraron diferencias significativas
en la contribución de las diferentes especies en relación con la distancia a
las fuentes de agua. El modelo de regresión no lineal fue el que mejor se
ajustó entre la fuente de agua y la receptividad ganadera para ambas unidades
geomorfológicas. La distancia a las fuentes de agua permanente es un buen
indicador de la presión ganadera acumulada en regiones semiáridas como el área
de estudio.
Palabras clave: Chaco árido,
pastizales naturales, piosfera, geomorfología
Originales: Recepción: 07/11/2023 - Aceptación: 26/03/2024
Introduction
Like
other arid and semi-arid regions, the Arid Chaco exhibits heterogeneous spatial
patterns of degradation driven by the uneven distribution of drinking throughs
(4, 19). The availability of
permanent water sources is critical for livestock production in these areas.
Consequently, a radial pattern of grazing intensity, known as ‘piosphere,’
develops around water sources. Analysis of piospheres allows to quantify the
effects of radial attenuation of a disturbance on the system’s condition (8).
Piospheres
are areas around water sources that suffer from heavy grazing and trampling,
making it difficult for vegetation to establish (degraded zones) (22).
Management practices near these water bodies often involve continuous grazing
without rest periods for the land, leading to changes in plant composition (10).
As a result, plant communities near wáter sources are dominated by annual
grasses and/or species with little grazing value (23).
Grassland
dynamics can be conceptualized using a state-and-transition model (STM), which
describes vegetation as existing in discrete states with transitions triggered
by natural events or management practices (5, 38).
Within an STM framework, each state represents a distinct plant community
reflecting the current ecological conditions relative to a climax community.
Transitions within a state are considered reversible, while transitions between
states can be irreversible depending on disturbance severity (26,
36).
In
interaction with livestock, geomorphology shapes the spatial patterns and
dynamics of plant communities (27).
In arid and semi-arid environments, water availability is the primary control
of vegetation structure and function, surpassing even the influence of the
physical and chemical characteristics of each geomorphological unit (24).
Recent sedimentary environments, are particularly sensitive to the colonization
strategies employed by vegetation given the specific arrangement and structures
of their deposits (15).
Evaluating
the impact of grazing on ecosystem integrity becomes difficult, especially in
systems where the original condition is unknown given widespread and
irreversible transformations of plant communities (10).
Furthermore, when a system has surpassed a critical threshold and transitioned
to an alternative stable state, experiments utilizing grazing exclusion methods
may yield misleading interpretations (9).
An
alternative approach to evaluate grazing is to interpret current vegetation
assemblages in the context of accumulated livestock pressure and associated
management practices. Proxy indicators such as proximity to water sources (1),
livestock posts (32),
and even specific plant growth forms or species abundance (13,
18) can be employed for this purpose. These approaches often provide
solid information to inform management decisions, particularly when time
constraints prevent reliance on long-term grazing trials (10).
This study
investigates the impact of extensive livestock farming on botanical composition
and livestock receptivity within distinct geomorphological units of the Arid
Chaco region in San Juan, Argentina. We aim to contribute to the understanding
of the ecological state of plant communities in semi-arid areas assessing
livestock receptivity potential and informing sustainable management
strategies. The study area exhibits diverse plant communities with varying
physiognomies, shaped by the combined effects of livestock farming, forestry exploitation,
and interactions with geomorphological processes.
Within this
framework, our study hypothesizes that botanical composition within each
geomorphological unit varies in response to distance from water sources. This
prediction is based on permanent water sources negatively influencing the
ecological states of plant communities, leading to differential livestock
receptivity.
Materials
and methods
Study
area
The study area
covers approximately 1000 km2 within the Valle Fértil department,
located between parallels 30°50’ and 30°29’ S and meridians 67°27’ and 67°12’
W. It is located east of the Sierras de Valle Fértil-La Huerta within the Bajo
Oriental depression, bordering La Rioja to the east. This region represents a
unique and characteristic expression of the Arid Chaco in San Juan, exhibiting
distinctive physiographic, climatic, social, and productive features. Its
climate is classified as arid (BWk) according to the Köppen-Geiger system (20), with an
average annual temperature of 17.9°C. Precipitation falls within the 200-300 mm
isohyets. The vegetation comprises an open forest dominated by Aspidosperma
quebracho blanco, Neltuma flexuosa, and Bulnesia retama. A
shrub layer rich in Larrea divaricata is present, while the herbaceous
layer is well-represented by genera such as Leptochloa, Setaria, Aristida,
and Pappophorum, among others.
Basin
delimitation
A
high-resolution (12.5-meter pixel size) Alos Palsar Digital Elevation Model
(DEM) was used to delineate the basins. Flow direction derived from the DEM was
employed to identify the basins associated with the main channels within the
study area. GRASS GIS software, a free and open-source module linked to QGIS
3.14.15 (30), was used for
the digital processing of the DEM.
Given the study
area corresponds to a plain, vectors representing the mountain range were
excluded from the analysis. Consequently, sampling was focused on the
geomorphological units of the foothills (Piedemonte) and the river floodplain
(Floodplain) (figure 1).

Fuente/Source: IGN (Instituto Geográfico Nacional,
República Argentina).
Figure 1. Location
of study sites on the Arid Chaco phytogeographic province of San Juan and La
Rioja, Argentina. Projection: Posgar 2007; Argentina/2; EPSG: 5344.
Figura 1. Ubicación
de los sitios de estudio en la Provincia Fitogeográfica del Chaco Árido de la
provincia de San Juan y La Rioja, Argentina. Proyección: Posgar 2007;
Argentina/2; EPSG: 5344.
Location
of livestock posts
The study area
encompasses a diversity of fenced enclosures with varying surface areas. Each
enclosure has a water source (dam, well, or drinker) for livestock located in
the vicinity of the post. All sampled fields employed a continuous grazing
regime, where cattle roam freely with unrestricted access to a central water
source. During periods of extreme drought, ranchers are forced to either sell
animals at a reduced price or pay to graze them in fields with superior forage
availability.
This study
adopts the methodology proposed by Cingolani et
al. (2008),
who emphasizes the value of distance to permanent water sources as an
objective, measurable, and precise indicator for assessing the long-term
effects of extensive grazing.
Establishment
of distance ranges
Given the
extensive grazing behavior of livestock in the Arid Chaco region (16), we
categorized sampling locations according to their distance to water sources:
close range (0-1000 m), intermediate range (1001-2000 m), and far range
(>2001 m). This approach was applied within each geomorphological unit
(Piedemonte and Floodplain). Thirty 50-meter transects were established within
each unit, with readings taken at 50 cm intervals along the transects (figure
1).
In the Piedemonte, ten transects were located close to water sources (within
1000 m, designated PMC), ten at intermediate distances (1001-2000 m, PMI), and
ten at a far range (>2001 m, PML). Similarly, in the Floodplain, ten
transects were placed near water sources (<1000 m, PLC), ten at intermediate
distances (1001-2000 m, PLI), and ten at a far range (>2001 m, PLL) (figure
1).
Determination
of botanical composition
Botanical
composition was quantified at peak growing season using the modified Point
Quadrat method (28) in April and
May, following the rainy season. The resulting data were then used to calculate
percent coverage and specific contribution per contact (CSC) for each species.
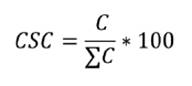
where:
C
= Contacts of a species
ΣC=
Sum of the contacts of all species
Determination
of field forage receptivity for bovine cattle
Livestock receptivity
refers to the amount of forage required to sustain an Equivalent Cow Unit (EV).
An EV is defined as a 400 kg cow that gestates and raises a calf to 6 months
old (weighing 160 kg), including the forage consumed by the calf (Passera,
C.P.Q.). One EV can be supported by 100 Pastoral Value (VP) units.
Plant species
for calculating Pastoral Value (VP) were selected based on established criteria
for determining specific quality indices (19, 29). Table
1,
presents their classification according to livestock preference: Preferential
(P) - readily consumed species without selection; Good (G) - species initially
rejected compared to Preferential ones; Regular (R) - species presenting some
difficulty in consumption.
Table
1. Contribution of each species, total
coverage, and diversity indices in each sector considered for sampling within
each Geomorphological Unit separately.
Tabla 1.
Aporte de cada especie, cobertura total e índices de diversidad en cada uno de
los sectores considerados para el muestreo en cada Unidad Geomorfológica por
separado.
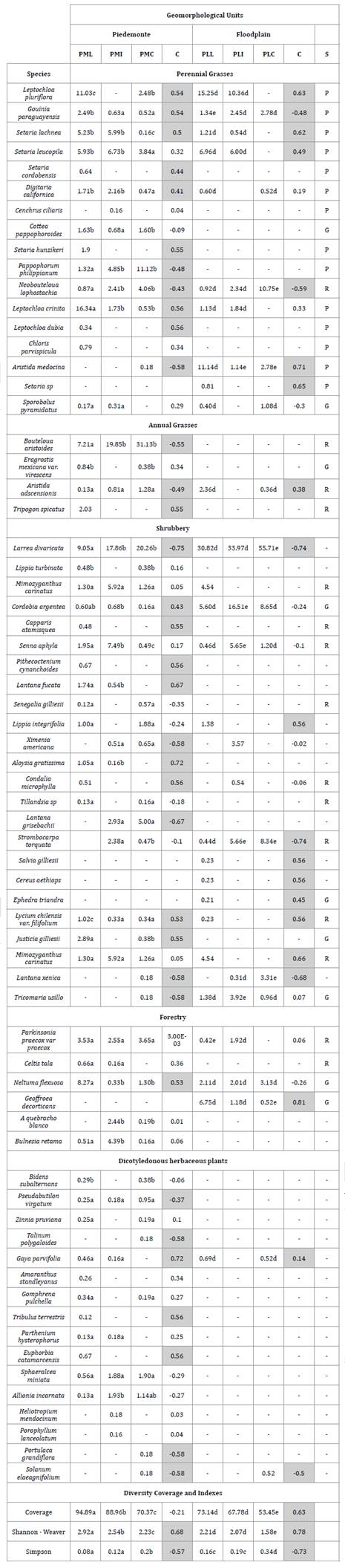
Different letters indicate significant differences
(P < 0.05). The column labeled C represents the correlation of each species,
total coverage, and diversity indices with respect to the distance from water
sources. Significant correlations (P < 0.05) are indicated by grayed cells.
Letras diferentes indican diferencias significativas
(P<0,05). La columna indicada con C representa la correlación de cada
especie, cobertura total e índices de diversidad, respecto de la distancia con
las fuentes de agua. Las correlaciones significativas (P<0,05) se indican
con los casilleros pintados de gris.
The Pastoral
Value (VP) is calculated by the following formula:
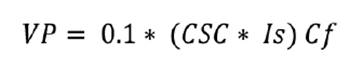
where:
0.1
= constant coefficient
CSC
= Specific Contribution per Contact
Cf
= Forage Coverage
Is
(Specific Quality Index) = ranging from 1
to 10, representing the classification of species based on their suitability
and potential as forage.
Statistical
analysis
Variation in
botanical composition across different areas was explored using principal
component analysis (PCA) based on a correlation matrix among all present
species and their respective contact-specific contributions (CSC) within the
community.
The influence of
distance to water sources on botanical composition, diversity (Shannon-Weaver
and Simpson), and total cover was assessed using analysis of variance (ANOVA)
with a completely randomized design. Separate ANOVAs were conducted for each
geomorphological unit (Piedemonte and Floodplain), treating distance categories
(PMC, PMI, PML and PLC, PLI, PLL) as fixed effects. Additionally, a Pearson
correlation analysis was performed to explore relationships between plant
species, diversity indices, and total coverage. Data were square root
transformed to address non-homoscedasticity and normality violations. Means
were compared using Tukey’s test. Data results are presented using
untransformed means for ease of interpretation.
Visual
exploration of the data involved plotting livestock receptivity against
distance to water sources for each geomorphological unit. This analysis
revealed a negative correlation up to a certain distance, followed by a plateau
effect. Consequently, non-linear models were fitted for each unit to account
for this observed pattern (12).
Results
Botanical
composition
A total of 26
families were recorded, with Poaceae being the best-represented family,
comprising 22 species. Fabaceae, Verbenaceae, and Asteraceae followed
in abundance, with 7, 6, and 4 species, respectively.
The Piedemonte
plant community is dominated by Leptochloa crinita. Additionally, Leptochloa
pluriflora, Bouteloua aristoides, Setaria leucopila, Setaria
lachnea, and Gouinia paraguayensis are present in PML sectors. In
PMI and PMC sectors, the Bouteloua aristoides contribution increases,
while the contributions of other grass species (except Setaria leucopila and
Setaria lachnea, which maintain their presence in the PMI sector) decrease.
The Piedemonte shrub layer is dominated by Larrea divaricata,
accompanied by Mimosiganthus carinatus, and Senna aphylla. The
forest layer exhibits a distinct zonation: Prosopis flexuosa dominates
in PML sectors, Bulnesia retama in PMI sectors, and Aspidosperma
quebracho blanco occurs at low densities across all sectors.
Within the
Floodplain, shrub communities dominate the vegetation structure. Larrea
divaricata is the most abundant species, with accompanying species Cordobia
argentea and Mimozyganthus carinatus in PLL sectors. In PLI sectors,
shrub species with increased contributions include Cordobia argentea, Senna
aphyla, and Prosopis torquata. Similarly, Larrea divaricata shows
a significant increase in contribution within PLC sectors, accompanied by Prosopis
torquata, Lantana xenica, and Cordobia argentea.
Following the
shrub layer, grasses contribute significantly to the Floodplain community. Aristida
mendocina and Leptochloa pluriflora are the most prominent grasses,
with Setaria leucopila present to a lesser extent in PLL sectors.
However, in PLC sectors, Leptochloa pluriflora disappears, and Aristida
mendocina abundance declines significantly. Conversely, Neobouteloua
lophostachia becomes the dominant grass species.
The forest layer
plays a minor role in the Floodplain, with Geoffroea decorticans being
the most prevalent species in PLL sectors, while Prosopis flexuosa dominates
in PLI and PLC sectors (table 1).
Across both
geomorphological units, total coverage and the Shannon-Weaver diversity index
exhibit a significant rise with increasing distance from water sources.
Conversely, Simpson’s dominance index shows a significant positive correlation
with proximity to water sources (table 1).
Principal
Component Analysis (PCA) revealed a variation pattern in botanical composition
across the sampled geomorphological units. The first two principal components
explained 61% of the total species variance, effectively separating sectors
within the Floodplain from those in the Piedmont. Within the Piedmont, sectors
located close and halfway to water sources (PMC and PMI) were distinct from
those situated further away (PML) (figure 2).
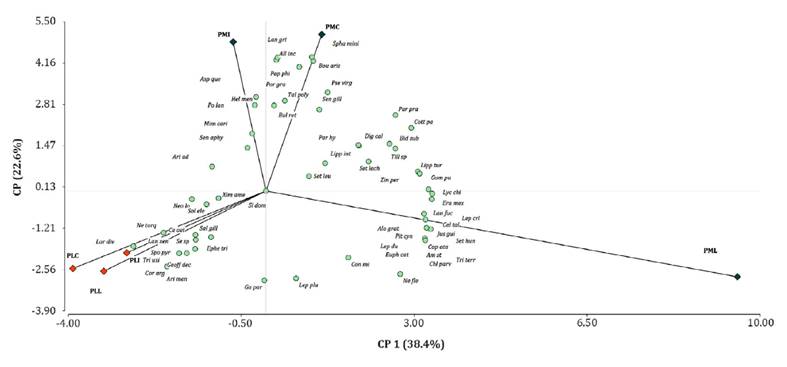
Figure 2. Principal
Component Analysis (PCA) ordination diagram of sampling sectors (PMC, PMI, PML,
PLC, PLI, PLL) and associated plant species.
Figura 2. Diagrama
de la ordenación del Análisis de Componentes Principales (ACP) de los sectores
seleccionados para los muestreos (PMC, PMI, PML, PLC, PLI, PLL) y las especies
asociadas.
Within the
alluvial plain, CP1 and CP2 displayed the most negative values for L.
divaricata (Lar div), C. argentea (Cor arg), T. usillo (Tri
usi), and G. paraguayensis (Ga par). In contrast, within the floodplain
and specifically PML sectors, CP1 exhibited the most positive values for J.
gilliesii (Jus gui), L. chilensis var. filifolium
(Lyc chi), S. cordobensis (Set cor), L. crinita (Lep cri), C.
tala (Cel tal), N. flexuosa (Ne fle), and L. pluriflora (Lep
plu). Similarly, positive values in CP2 were associated with PMI and PMC
sectors for B. aristoides (Bou aris), A. quebracho blanco (Asp
que), S. miniata (Spha mini), A. incarnata (All inc), L.
grisebachii (Lan gri), P. philippianum (Pap phi), H. mendocinum (Hel
men), and P. lanceolatum (Po lan) (figure 2).
Livestock
receptivity and distance to water sources
Non-linear
regression analysis proved the best fit between livestock receptivity (Ha.EV-1)
and distance to water sources (DP), for both the Piedemonte (PM) and Floodplain
(PL) units (figure 3).
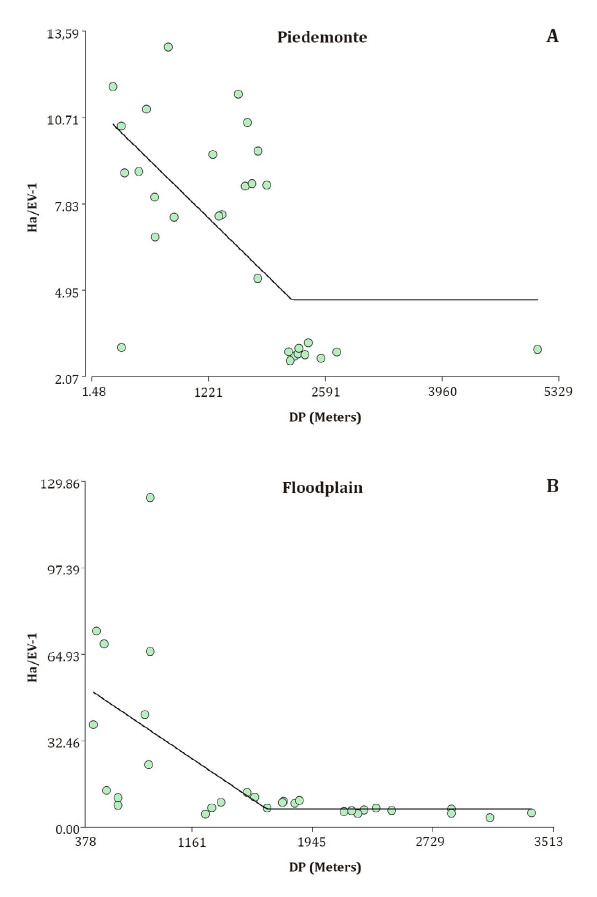
Figure 3. Non-linear
regression model between livestock receptivity (Ha.EV-1) and
distance to water source (DP) in the Piedemonte (A) and Floodplain (B).
Figura 3. Modelo
de regresión no lineal entre la receptividad ganadera (Ha.EV-1) y la
distancia a la fuente de agua (DP) en el Piedemonte (A) y en la Planicie de
inundación (B).
Discussion
Our findings
reveal changes in floristic composition across both geomorphological units,
with distance to water sources playing a significant role (figure 2 and table
1).
The study area is characterized by fenced fields of varying sizes and shapes,
along with an irregular distribution of watering holes, dams, and/or livestock
shelters. This heterogeneity contributes to a highly diverse botanical
composition within plant communities. Szymañski et
al. (2022)
attribute similar variations primarily to livestock management practices.
Specifically, our results indicate the presence of plant communities dominated
by perennial forage grasses and others dominated by non-forage annual grasses.
In such cases, the potential for transition towards improved ecological
conditions exists due to the presence of seed sources from desirable species,
facilitating the natural rehabilitation of nearby degraded areas (10).
The results
align with established patterns of overgrazing documented by Briske
et al. (2006),
where plant communities near water sources (PMC and PLC) are dominated by anual
grasses and/or non-forage species (table 1). While grazing
responses vary among species, when grazing pressure alters biotic structures
and interactions changes in species coverage can be attributed to underlying
biotic mechanisms of grazing impact. Several studies support this notion,
demonstrating that grazing pressure favors plant traits associated with rapid
growth, regeneration, annual life cycles, and a ruderal strategy (11,
14, 32).
Across the
geomorphological units, perennial grasses exhibited a significant trend of
increased colonization away from water sources (table 1). Conversely,
annual grasses and species with low forage value showed the opposite pattern.
This aligns with established knowledge on grazing pressure inducing species
turnover, favoring some species while hindering others (10). Briske
et al. (2006),
further suggests that heterogeneous livestock use is common in homogeneous
landscapes (i.e., within a single geomorphological unit). This
heterogeneity contributes to an increase in landscape physiognomic diversity.
However, increased livestock density can lead to a decline in unused areas (37). While
maintaining some level of unused sites is important for overall community
coexistence (10), excessively
large paddocks with few water sources or shelters may allow even high livestock
densities to coexist with unused areas (21, 26).
Total coverage
and Shannon-Weaver diversity in the geomorphological units exhibited a
significant decrease near water sources (table 1). Conversely,
Simpson’s dominance index shows a positive trend in these areas. This pattern
aligns with observations in other semi-arid African grasslands, where
grazing-sensitive forage species are replaced by those more resistant to
grazing. However, this replacement may not fully compensate for diversity
losses, potentially leading to environments dominated by a few resilient
species. In this context, grazing regimes exceeding the historical range
experienced by the ecosystem are likely to induce a significant decline in
overall diversity (16).
The observed
variations in floristic composition may be explained by mechanisms of
resilience to harsh conditions proposed by Cingolani et
al. (2008)
for arid and semi-arid grasslands. These mechanisms, such as reduced sprouting
and production of long-lived seeds, can confer some resistance to short periods
of intense grazing. However, this resilience is insufficient to withstand
continuous grazing for extended periods, particularly under moderate to high
grazing pressure. A study evaluating intensity versus grazing strategies in
perennial forage grasses of the Arid Chaco region found that while increased
defoliation intensity may maximize short-term biomass yield, it negatively
impacts long-term sustainability and produces detrimental residual effects on
grasslands (31). Importantly,
the study identified continuous grazing combined with high defoliation
intensity as the least sustainable management practice for this ecosystem (31).
In these
systems, continuous grazing by introduced herbivores without proper stocking
management is highly likely to cause widespread extinctions and/or significant
declines of specific plant species (10, 21, 25). However, a
contrasting recovery trajectory may emerge when degradation and/or extinction
of desirable species occurs homogenously across large areas (several square
kilometers) and grazing pressure is subsequently relieved (10,
21).
An alternative stability perspective for degraded ecosystems emphasizes
persistence relative to the livelihood needs of ecosystem users. If the
timeframe for ecosystem recovery surpasses persistent from a practical
viewpoint. Therefore, a ‘persistent decline’ of an ecosystem service may not
always be associated with a critical threshold and a shift to a new ecological
state. Slow recovery within a single stability domain can also lead to
persistent declines (23).
Livestock
receptivity values in this study range from 47.02 to 2.86 Ha.EV-1 (figure
3).
These values are consistent with observations from other arid and semi-arid
regions in Patagonia (23), Puna, Monte,
and the driest parts of the Arid Chaco, where reported livestock receptivities
typically exceed 6 Ha.EV-1 (3). Nevertheless,
the value of 8.92 Ha.EV-1 measured in the PMC sector deviates from
historical data for degraded foothills, which reported values around 24 Ha.EV-1
(33). This discrepancy may be attributed to
interannual precipitation variations, known to cause substantial differences
(>300%) in grassland productivity within degraded areas of the Arid Chaco (11).
Despite using
the same model for both geomorphological units, the estimated values and rates
of change differed (figure 3). These
non-linear relationships align with the findings of Sasaki
et al. (2008),
who highlight the non-linear nature of ecological patterns and processes in
response to grazing pressure. Furthermore, the estimated values from our models
are consistent with other studies conducted in the region (2,
7, 11).
Additionally, the observed spatial patterns of vegetation change correspond to
documented livestock grazing behavior in the Arid Chaco (16). This
distribution pattern of livestock receptivity informs spatial dynamics within
grazing plots, allowing for the establishment of criteria for plot size and
water source distribution.
Conclusions
Across both
geomorphological units, distance to water sources significantly influences
floristic composition, which in turn affects livestock receptivity. Areas
closer to water sources are dominated by annual grasses and/or non-forage
species, resulting in lower livestock receptivity. Conversely, areas farther
from water sources are dominated by perennial grasses with higher forage value,
leading to increased livestock receptivity. A long history of domestic grazing
has shaped the heterogeneity of plant communities, influencing species
diversity and dominance. This translates to a landscape with varying ecological
conditions (good, fair, and bad) distributed in different proportions.
Quantifying the extent of each condition is crucial for optimizing water source
distribution and achieving biologically and economically sustainable grazing
management strategies. Notably, non-linear models provided the best fit between
distance to water sources and livestock receptivity for both geomorphological
units under study (figure 3).
Acknowledgements
We
thank the anonymous reviewers and English Professor Jimena Olivares for their
comments on this article. This research is supported by INTA - EEA Pocito and
CONICET - CCT San Juan. Argentina.
1. Adler, P. B.;
Milchunas, D. G.; Sala, O. E.; Burke, I. C.; Lauenroth, W. K. 2005. Plant
traits and ecosystem effects of grazing: comparison of the American sagebrush
steppe and the Patagonian steppe. Ecological Applications. 15 (2): 774-792.
2. Blanco, L.
J.; Ferrando, C. A.; Biurrun, F. N. 2009. Detección remota de patrones
espaciales y temporales de vegetación en dos sistemas de pastoreo. Ecología
y manejo de pastizales. 62(5): 445-451.
3. Blanco, L.
J.; Durante, M.; Ferrante, D.; Quiroga, R. E.; Demaria, M. R.; Di Bella, C. M.
2019. Red nacional de monitoreo de pastizales naturales de Argentina:
productividad forrajera de la vegetación extrapampeana. Revista de
Investigaciones Agropecuarias. 45(1): 89-108.
4. Blanco, L.
B.; Aguilera, M. O.; Paruelo, J. M.; Biurrum, F. N. 2008. Grazing effect on
NDVI acros san aridity gradient in Argentina. Journal of Arid Environments. 72:
764-776.
5. Briske, D. D.
2017. Rangeland systems processes, management and challenges. Springer.
10.1007/978-3-319-46709-2
6. Briske, D.
D.; Fuhlendorf, S. D.; Smeins, F. E. 2006. A unified framework for assessment
and application of ecological thresholds. Rangeland Ecology and Management. 59:
225-236.
7. Calella, H.
F.; Corzo, R. R.; Gómez, J. C.; Reynoso, A. A.; Zalazar, A.; Murúa, S.;
Ricarte, A. 2006. El Chaco árido de La Rioja: vegetación y suelos. Pastizales
naturales. Ediciones INTA. Buenos Aires. Argentina.
8. Chillo, V.;
Ojeda, R. A. 2014. Disentangling ecosystem responses to livestock grazing in
drylands. Agriculture, Ecosystems and the Environment. 197: 271-277.
9. Cingolani, A.
M.; Noy-Meir, I.; Diaz, S. 2005. Grazing effects on rangeland diversity:
diversity-intensity and state and transition models. Ecological Applications.
15: 757-773.
10. Cingolani,
A. M.; Renison, D.; Tecco, P. A.; Gurvich, D. E.; Cabido, M. 2008. Predicting
cover types in a mountain range with long evolutionary grazing history: a GIS
approach. Journal of Biogeography. 35(3): 538-551.
11. Díaz R., O.
2007. Utilización de pastizales naturales. Encuentro Grupo Editor. Córdoba,
Argentina.
12. Di Rienzo,
J. A.; Casanoves, F.; Balzarini, M. G.; Gonzalez, L.; Tablada, M.; Robledo, C.
W. 2020. InfoStat versión 2020. Centro de Transferencia InfoStat, FCA.
Universidad Nacional de Córdoba. Córdoba. Argentina. http://www.infostat.com.ar
13. Dyksterhuis,
E. J. 1949. Condition and management of rangeland based on quantitative
ecology. Journal of Range Management. 41: 450-459.
14. Espinoza, J.
J. O.; Ayala, C. C.; Castillón, E. E.; Saldivar, F. G.; Sauceda, J. U.; Jurado,
E.; Hernández, E. O. 2017. Livestock effect on floristic composition and
vegetation structure of two desert scrublands in northwest Coahuila, Mexico.
The southwestern naturalist. 62(2): 138-145.
15. Flores, D.
G.; Suvires, G.; Dalmasso, A. 2015. El análisis geomorfológico como base para
el estudio de la vegetación nativa: Sierra Chica de Zonda, Precordillera
Oriental de Argentina. Cuadernos de Investigación Geográfica. 41(2): 427-444.
16. Hanke, W.;
Böhner, J.; Dreber, N.; Jürgens, N.; Schmiedel, U.; Wesuls, D.; Dengler, J;
2014. The impact of livestock grazing on plant diversity: an analysis across
dryland ecosystems and scales in southern Africa. Ecological Applications.
24(5): 1188-1203.
17. Herrera
Conegliano, O. A. 2018. Comportamiento en pastoreo del ganado bovino Criollo
Argentino y Aberdeen Angus ecotipo riojano, en pastizales naturales del Chaco
Árido. Doctoral dissertation, Facultad de Ciencias Agrarias. Universidad
Nacional de Mar del Plata.
18. Jauffret,
S.; Lavorel, S. 2003. Are plant functional types relevant to describe
degradation in arid, southern Tunisian steppes? Journal of Vegetation Science.
14(3): 399-408.
19. Karlin, M.
S. 2013. Relaciones suelo-planta en el ecosistema Salinas Grandes, Provincia de
Catamarca (Argentina). Doctoral thesis. Facultad de Ciencias Agropecuarias,
Universidad Nacional de Córdoba.
20. Koppen, W.
1936. Das geographische System de Klimate. Handbuch der klimatologie.
Borntraeger. Berlín. Bd. 1.C.
21. Landsberg,
J.; James, C. D.; Morton, S. R.; Müller, W. J.; Stol, J. 2003. Abundance and
composition of plant species along grazing gradients in Australian grasslands.
Journal of Applied Ecology. 40(6): 1008-1024.
22. Magliano, P.
N.; Breshears, D. D.; Murray, F.; Niborski, M. J.; Nosetto, M. D.; Zou, C. B.;
Jobbágy, E. G. 2023. South American Dry Chaco rangelands: Positive effects of cattle
trampling and transit on ecohydrological functioning. Ecological Applications.
33(3): e2800.
23. Merdas, S.;
Kouba, Y.; Mostephaoui, T.; Farhi, Y.; Chenchouni, H. 2021. Livestock
grazing-induced large-scale biotic homogenization in arid Mediterranean steppe
rangelands. Land Degradation & Development. 32(17): 5099-5107.
24. Monger, C.;
Bestelmeyer, T. 2006. The soil-geomorphic template and biotic change in arid
and semiarid ecosystems. Journal of Arid Environments. 65: 207-218.
25. Oliva, G.;
Paredes, P.; Ferrante, D.; Cepeda, C.; Rabinovich, J. 2019. Remotely sensed
primary productivity shows that domestic and native herbivores combined are
overgrazing Patagonia. Journal of Applied Ecology. 56(7): 1575-1584.
26. Oñatibia, G.
R. 2021. Grazing management and provision of ecosystem services in Patagonian
arid rangelands. In Ecosystem Services in Patagonia: A Multi-Criteria Approach
for an Integrated Assessment. Springer International Publishing. p. 47-74.
27. Parker, K.
C.; Bendix, J. 1996. Landscape-Scale geomorphic Influences on vegetation
patterns in four environments. Physical Geography. 17(2): 113-141.
28. Passera, C.
B.; Dalmasso, A. D.; Borsetto, O. 1983. Método de “point quadrat modificado”.
Taller de arbustos forrajeros para zonas áridas y semiáridas. 71-79.
29. Passera C.
B.; Borseto O. 1986. Determinación del “Índice de Calidad Específico”. Actas
del Taller de Arbustos Forrajeros. Grupo Regional FAO-IADIZA, Mendoza.
30. QGIS, Equipo
de desarrollo. 2020. Sistema de información geográfica de código abierto.
Fundación Fuente Geoespacial. Versión 2.18.20. https://qgis.org/es/site
31. Quiroga, R.
E.; Blanco, L. J.; Namur, P. R. 2018. Defoliation intensity and effects of
simulated grazing strategies in three C4 grasslands. Range Ecology and
Management. 71(1): 58-66.
32. Sasaki, T.;
Okayasu, T.; Jamsran, U.; Takeuchi, K. 2008. Threshold changes in vegetation
along a grazing gradient in Mongolian rangelands. Journal of Ecology. 96(1):
145-154.
33. Scaglia, J.
A.; Flores, D. G.; Martinelli, M. 2021. Productividad de los pastizales
naturales en diferentes unidades geomorfológicas de las Sierras Pampeanas de
Argentina. Ecosistemas. 30(2): 2104-2104.
34. Skarpe, C.
2000. Desertification, no-change or alternative states: Can we trust simple
models on livestock impact in dry rangelands? Applied Vegetation Science. 3(2):
261-268.
35. Szymañski,
C.; Villagra, P. E.; Aschero, V.; & Alvarez, J. A. 2022. Interactive
effects of chronic anthropogenic disturbances on Prosopis forest structure in
Monte Central, Argentina. Southern Ecology. 32(1): 108-121. https://doi.org/10.25260/EA.22.32.1.0.1800
36.
Vignoni, A. P.; Peralta, I. E.; Abraham, E. M. 2023. Fragmented areas due to
agricultural activity: native vegetation dynamics at crop interface
(Montecaseros, Mendoza, Argentina). Revista de la Facultad de Ciencias
Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 55(2): 46-60. DOI:
https://doi.org/10.48162/rev.39.108
37.
Weber, K. T.; Horst, S. 2011. Desertification and livestock grazing: the role
of sedentarization, mobility and rest. Pastoralism: Research. Policy and
Practice. 1(1): 1-11.
38.
Westoby, M.; Walker, B.; Noy-Meir, I. 1989. Opportunistic management of
grasslands that are not in equilibrium. Range Ecology and Management/Journal of
Range Management Archives. 42(4): 266-274.