Revista de la Facultad de Ciencias
Agrarias. Universidad Nacional de Cuyo. En prensa. ISSN (en línea) 1853-8665.
Original article
Radial
Growth Dynamics and Drought Resilience in Pinus pinea L. Plantations
from Central-Western Argentina: Implications for Forestry Development
Dinámica
del Crecimiento radial y resiliencia a la sequía de Pinus pinea L. en
plantaciones del centro-oeste argentino: implicancias forestales
Luciano Diaz
Dentoni3,
1Instituto Argentino de Nivología, Glaciología y Ciencias
Ambientales. Consejo Nacional de Investigaciones Científicas y Técnicas
(IANIGLA CONICET). Laboratorio de Dendrocronología e Historia Ambiental. Av.
Dr. Adrian Ruiz Leal. Parque Gral. San Martín. M5500 Mendoza. Argentina.
2Universidad Nacional de Cuyo. Facultad de Ciencias Agrarias.
Cátedra de Economía y Política Agraria. Almirante Brown 500. M5528AHB. Chacras
de Coria. Mendoza. Argentina.
3Universidad Nacional de Cuyo. Facultad de Ciencias Agrarias.
Almirante Brown 500. M5528AHB. Chacras de Coria. Mendoza. Argentina.
4Universidad Mayor. Facultad de Ciencias. Hémera Centro de
Observación de la Tierra. Camino La Pirámide 5750. Huechuraba. Santiago.Chile.
*spiraino@fca.uncu.edu.ar
Abstract
Forests play a
crucial role in ecological stability, carbon sequestration, habitat provision and economy. As climate change intensifies, increasing
drought frequency and severity challenge our understanding of forest
resilience. Based on this premise, we examined radial growth dynamics and
drought response of Pinus pinea L. in Mendoza Province in both mesic and
xeric conditions. Using dendrochronological techniques, we assessed the long
and short-term effects of soil and atmospheric drought on radial growth trends
at two irrigated plantations with contrasting environments. Growth dynamics
reflected differences in soil, climate, and irrigation. Growth rates were
significantly higher at the mesic stand, which received nearly twice the
precipitation and irrigation compared to xeric one. In contrast, growth at the
xeric site was strongly limited by early-summer atmospheric drought, while
late-growing season soil moisture and climatic conditions affected tree-ring
development at the mesic site. Growth resilience to extreme events experienced
site dependence, with edaphic drought exerting a stronger negative effect than
atmospheric dry spells at the mesic stand. Our results underscore the
importance of integrating short- and long-term drought assessment into P.
pinea management strategies and support the potential of stone pine
plantations in extra-Mediterranean South America for sustainable forestry under
changing climatic conditions.
Keywords: climate change,
forest management, mendoza, tree-rings
Resumen
Los bosques son
esenciales en la estabilidad ecológica, la captura de carbono, la provisión de
hábitats y de recursos económicos. Ante el aumento de la frecuencia y severidad
de las sequías asociado al cambio climático,
comprender la resiliencia forestal resulta crucial. Este estudio analizó
mediante métodos dendrocronológicos los efectos a corto y largo plazo de las
sequías edáficas y atmosféricas sobre el crecimiento radial de Pinus pinea L.
en dos plantaciones irrigadas bajo condiciones ambientales contrastantes en el
centro-oeste argentino. Los patrones de crecimiento reflejaron diferencias
ambientales y de manejo. Las tasas de crecimiento fueron significativamente
mayores en el rodal mésico, que recibió casi el doble de precipitación e
irrigación que el xérico. En este último, el crecimiento se vio limitado por
sequías atmosféricas de comienzo del verano, mientras que en el mésico
influyeron las sequías edáficas y climáticas al final de la temporada de
crecimiento. La resiliencia ante eventos extremos mostró dependencia del sitio,
con un efecto negativo más marcado de la sequía edáfica en el rodal mésico. Los
resultados destacan la necesidad de integrar evaluaciones multitemporales de
sequía en el manejo de P. pinea y su potencial para un desarrollo forestal
sostenible en regiones extra-Mediterráneas de Sudamérica.
Palabras clave: cambio climático,
manejo forestal, Mendoza, anillos de crecimiento de árboles
Originales: Recepción: 20/05/2025 - Aceptación: 21/10/2025
Introduction
Forests maintain ecological
balance by storing carbon, regulating water cycles, and providing habitat for
numerous species (Barnes et al., 1997).
Recent climatic shifts have increased the focus on the resilience of natural
and planted forests under abiotic disturbances (Johnstone
et al., 2016). Drought threatens forest health and productivity, as
dry spells become more frequent and intense (IPCC, 2023).
Extreme drought events have severely impacted forests, reducing growth rates
and increasing tree mortality (Allen et al., 2010).
Due to their
genetic uniformity, plantation forests may respond differently to dry spells
than natural woodlands (Camarero et al., 2021;
Navarro-Cerrillo et al., 2023). Their capacity to withstand and
recover from a drought event dictates their long-term sustainability and
broader environmental and economic values. Even considering that forest
plantations only cover about 7% of the global forest area, they significantly
contribute to mitigating deforestation impacts on natural woodlands (FAO, 2020). This provides particular importance to the
in-depth understanding of their drought resilience.
Drought stress
manifests in two primary forms: atmospheric drought, driven by low humidity and
high evaporative demand, and edaphic drought, characterized by depleted soil
water (Mishra & Singh, 2010). These conditions
can occur separately or together, each uniquely impacting plant growth (Knutzen et al., 2017). Disentangling these
responses is key to predicting how trees will perform in a changing climate.
Forest responses to
extreme drought can be assessed by analyzing long- and short-term growth
trends. Long-term trends reveal climate-growth relationships, while short-term
responses measure resilience, that is, the capacity to withstand and recover
from dry spells (Lloret et al., 2011).
Dendrochronology provides a powerful method for investigating tree growth
responses to ecological variables, allowing us to reconstruct growth trends and
identify periods of stress, thereby illuminating species’ resilience and
adaptability (Piraino, 2020; Piraino et al.,
2022, 2024).
Pinus pinea L. (stone pine), a conifer prized for its edible seeds, holds
significant ecological and economic value (Mutke et
al., 2012). In its native Mediterranean range, the species now suffers
drought-related decline in cone yield (Calama et al.,
2020). Climate models project a contraction of its suitable habitat and a
negative trend in radial growth (Mechergui et al.,
2021; Natalini et al., 2024). Establishing extra-Mediterranean
plantations could help offset the impending global shortfall in nut production.
Growers have
successfully introduced stone pine to southern South America (Chile and
Argentina) (Muñoz et al., 2012), where it
adapts well to diverse conditions and promises economic benefits from
non-timber products (Loewe-Muñoz et al., 2012).
Recent studies have analyzed tree-ring variability and its relationship with
climate trends and extremes in Chilean plantations under typical Mediterranean
conditions (Loewe Muñoz et al., 2024ab). However,
no research has yet reconstructed radial growth dynamics in Argentina, where
the species thrives in semi-arid environments (Calderón et
al., 2008; Diaz Dentoni, 2024).
This study aims to
investigate how P. pinea radial growth interacts with drought
conditions in two irrigated plantations located in Argentina’s Mendoza
Province. We hypothesize that drought events significantly suppress growth,
with the stronger effects during extreme dry years. Our findings will clarify
the potential of this species for forestry in extra-Mediterranean regions,
especially as climate change threatens its native range.
Material
and Methods
Study
Site Characteristics
We selected two sites for this study: Dique Yaucha (hereafter
YAU; 34°00’03” S,
69°07’03” W) and Malargüe (hereafter MAL: 35°35’55” S, 69°31’37”
W), both located in southern Mendoza Province (figure 1).
These plantations represent the only undisturbed adult artificial forests in
the region (Piraino et al., 2021).
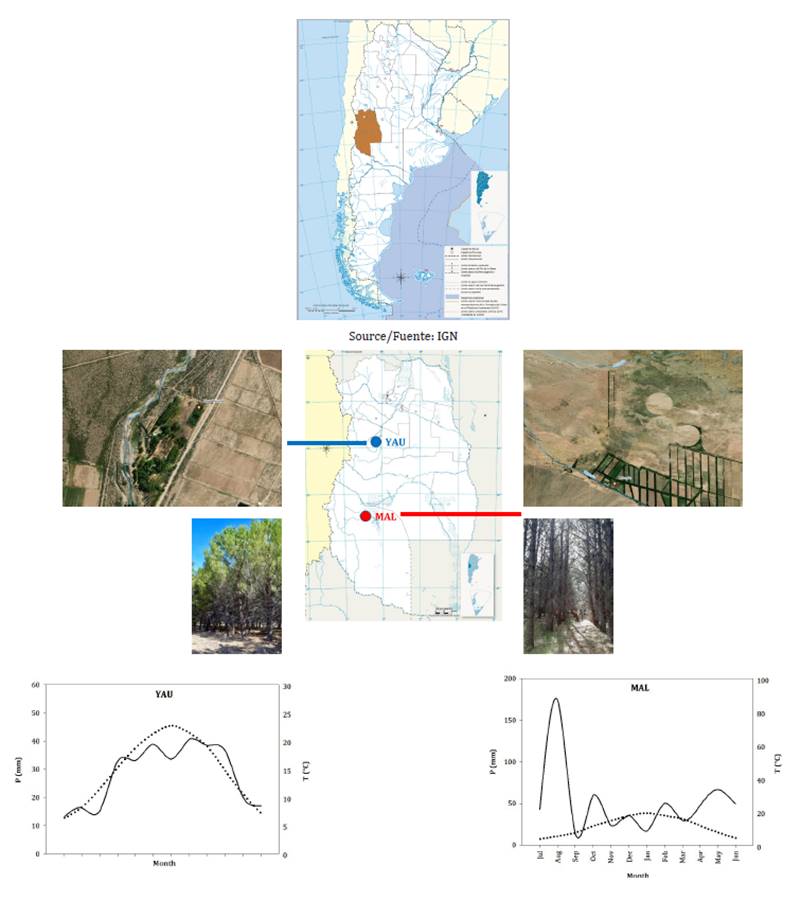
Precipitation
and temperature data belong to the INTA San Carlos (Dique Yaucha: 33.73°S,
69.1°W; http://siga.inta.gob.ar/#/), and Malargüe Aero (Malargüe: 35.48°S;
69.58°W; http://www.meteomanz.com/) gauge stations, covering the 2000-2020
period. T: monthly mean air temperature; P: monthly total rainfall. Gray dot
line refers to monthly mean air temperature, and black solid line to monthly
total rainfall.
Los
datos de precipitación y temperatura pertenecen a las estaciones climáticas
INTA San Carlos (Dique Yaucha: 33.73°S, 69.1°O; http:// siga.inta.gob.ar/#/) y
Malargüe Aero (Malargüe: 35.48°S; 69.58°O; http://www.meteomanz.com/), y cubren
el período 2000-2020. T: temperatura media mensual del aire; P: precipitación
total mensual. La línea de puntos grises indica la temperatura media mensual
del aire, y la línea negra continua, la precipitación total mensual.
Figure
1.
Geographical location of the sampled sites and Ombrothermic diagram drawn
according to the methods of Bagnouls
& Gaussen (1953).
Figura
1. Ubicación geográfica de los sitios muestreados y diagrama
ombrotérmico elaborado según los métodos de Bagnouls
& Gaussen (1953).
At both sites,
stone pine seedlings were planted between the late 1980s and early 1990s,
forming a monospecific plantation with a spacing of 3x3 m between rows and
trees. The plantations are situated at similar elevations (1213 m a. s. l. for
YAU vs 1470 m a. s. l. for MAL). The YAU site is characterized by semi-arid
conditions, with mean annual temperature of 15°C, and total annual
precipitation of 335 mm (figure 1). In contrast, MAL grows
under Mediterranean-type climate, with mean annual temperature and total annual
precipitation values of 12.6°C and 610 mm, respectively (figure 1).
Soils are gravelly to sandy with good permeability at YAU, and loam to clay
loam of alluvial origin at MAL. Both plantations receive supplementary
irrigation: approximately 300-350 mm annually at YAU and 500-600 mm at MAL,
evenly distributed across the irrigation season (September-July; current and
projected upper river water balance 2020-2021; G. Aguado, personal
communication). According to this characterization, we considered YAU and
MAL sites xeric and mesic, respectively.
Dendrochronological
Sampling and Tree-Ring Chronology Development
During the austral
spring of 2021 and 2022, we extracted one core per tree at breast height
(approximately 1.30 m above soil level) from 16 individuals at YAU and 10 at
MAL, respectively, by using a 5.15 mm increment borer. We selected sampled
trees based on their health status. Cores were glued onto wooden mounts, and
the transverse section was polished with progressively finer sandpaper to
highlight ring boundaries. Samples were scanned at 1200 dpi, and tree-ring
width (TRW) was measured to 0.001 mm precision with CooRecorder software (Maxwell & Larsson, 2021). Calendar years were
assigned following Schulman’s convention for the Southern Hemisphere (Schulman, 1956).
TRW series were
statistically validated with COFECHA software (Holmes,
1983). Two indices were calculated: 1) the mean correlation among
individual series (MC); and 2) mean sensitivity (MS), the relative year-to-year
variability in TRW, reflecting the species’ sensitivity to environmental
factors (Speer, 2010).
After statistical validation, individual chronologies were
standardized to remove the low-frequency, age-related trends and highlight the
high-frequency climate signal (Speer, 2010). Raw TRW
series were detrended using a 20-years spline with a 50% frequency cutoff
applied in ARSTAN40c software (Cook & Krusic, 2006). We used the
standardized residual chronology version for all drought-growth analyses to
minimize bias from non-climatic trends (Villalba &
Veblen, 1997).
Statistical
Analyses
To assess
differences in TRW trends among sites under varying environmental conditions,
we compared the raw ring-width site chronologies for the common period
1992-2020 with a Kruskal-Wallis test hosted in the InfoStat software (Di Rienzo et al., 2021). Then, we used site and
individual- standardized chronologies to evaluate the effect of drought on
growth at long and short-term timescales. For the long-term analysis,
correlation functions were calculated between the standardized
site-chronologies and two datasets: SPEI (Standardized Precipitation
Evaporation Index; Vicente-Serrano et al., 2010),
and soil moisture at 100 cm depth (SM100). Monthly values of both datasets were
obtained from the KNMI Climate Explorer for 1901-2018 (SPEI) and 1979-2016
(SM100) (Trouet & Van Oldenborgh, 2013; http://climexp.knmi.nl/). Correlation
functions were computed with DENDROCLIM2002 for the common period 1992-2016 (n
= 25) (Biondi & Waikul, 2004). We selected this
interval because the YAU chronology began in 1992 and SM100 data ended in 2016.
Based on prior research, stone pine plantations in South America (Loewe-Muñoz et al., 2022), a 13-month
time-window was selected, spanning June of the year preceding growth through June
of the growth year. No monthly irrigation data were available, which prevented
direct comparison between supplementary water inputs and ring development.
We assessed
short-term growth responses to extreme drought using the line of full
resilience (LFR; Schwarz et al., 2020). The
LFR approach evaluates the relationship between resistance and recovery sensu
Lloret et al., 2011; see below for
mathematical definitions. and compares it with the
theoretical scenario of full resilience (Schwarz et
al., 2020). This method provides an integrated assessment of tree
capacity to withstand drought stress (Schwarz et al.,
2020).
We defined drought
years based on the significant monthly windows identified by the correlation
function analysis, which we used to calculate annual series for SPEI and SM100.
We classified years with SPEI or SM100 values in the lowest 5th percentile as extreme
drought events. For these events, we calculated tree-level resistance (Rt)
and
recovery (Rc) indices as Rt = TRId/TRI_pred and Rc
= TRI_postd/TRId. TRI is the standardized
tree-ring index, d refers to the drought year, and pre and post
denote the year before and after extreme dry spell, respectively (Lloret et al., 2011). Short pre and
post time windows were chosen to minimize overlap among consecutive
drought events (see Results). Finally, Finally, we calculated the LFR as Rc = 1/Rt.
Results
We developed two
tree-ring chronologies from 10 (MAL) and 16 (YAU) wood samples (table
1 and figure 2). The chronologies covered 1991-2020 at
MAL and 1992-2021 at YAU (table 1). Mean annual raw TRW
(mTRW) was significantly higher at MAL than at YAU stand (H = 8.35; p =
0.0039; data not shown). Tree-ring statistics showed higher MC and MS values
for YAU (table 1).
Table 1.
Characteristics of the tree-ring chronologies.
Tabla
1. Características de las cronologías de anillos de crecimiento.
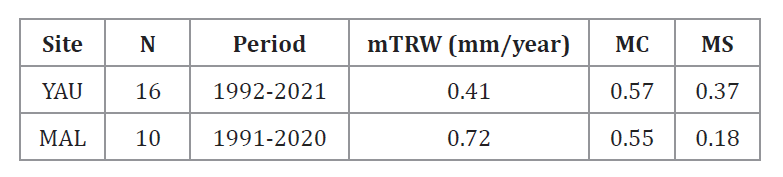
N:
Number of sampled trees per site. Period: time range of the sampled cores. mTRW: mean annual tree-ring width value. MC: mean
correlation between series at each stand. MS: mean sensitivity.
N:
Número de árboles muestreados por sitio. Period: rango temporal de las
muestras. mTRW: valor medio anual del ancho de anillo.
MC: correlación media entre series en cada rodal. MS: sensibilidad media.
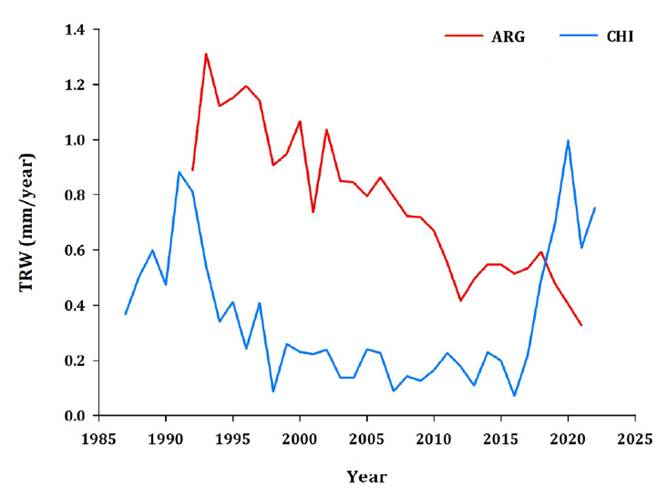
Figure
2.
Raw tree-ring width (TRW) site chronologies of the sampled stone pine
plantations.
Figura
2. Cronologías de datos brutos de ancho de anillo (TRW) de las
plantaciones de pino piñonero muestreadas.
Correlation function analyses
revealed that radial growth at both sites was significantly influenced by
long-term drought conditions, although responses differed between stands (figure 3). At MAL, standardized growth was positively related to
late spring (June; JUN) SPEI and SM100. At YAU, SPEI positively influenced ring
width during December (DEC) of the year of growth, while no significant relationship
emerged with SM100 (figure 3).
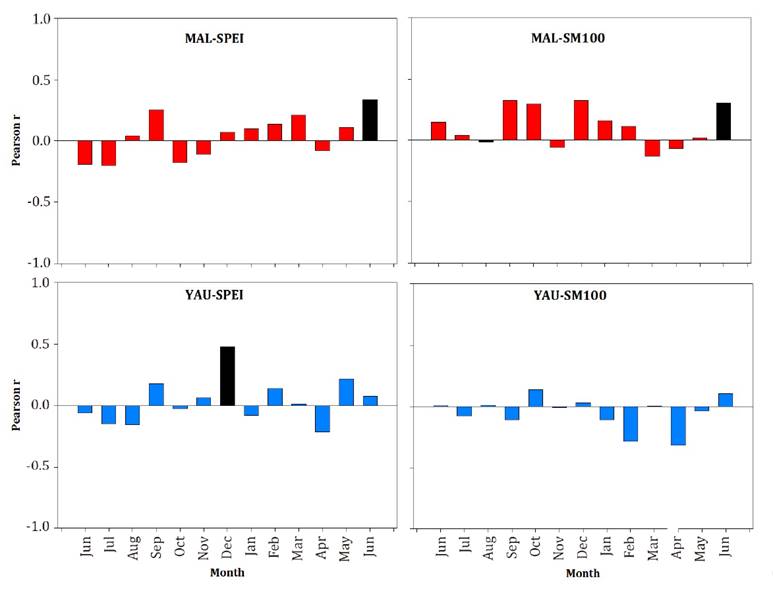
Black
bars refer to significant Pearson r values at p < 0.05 level.
Las
barras negras indican valores de correlación de Pearson significativos por p
< 0,05.
Figure
3.
Correlation functions results between each residual chronology and monthly SPEI
and SM100 values for the common period 1992-2016.
Figura
3. Resultados de las funciones de correlación entre cada
cronología residual y los valores mensuales de SPEI y SM100 para el período
común 1992-2016.
Based on these
results, we selected JUN-SM100 and JUN-SPEI to identify extreme drought events
at MAL, and DEC-SPEI at YAU stand. Statistical analyses identified three
extreme drought events at MAL (MAL-SM100 2010: MAL-SPEI 2012; MAL-SPEI 2014)
and two at YAU (YAU-SPEI 2003 and YAU -SPEI 2011) (figure 4).
We excluded the YAU -SPEI 2003 event, as trees were likely in their seedling
phase.
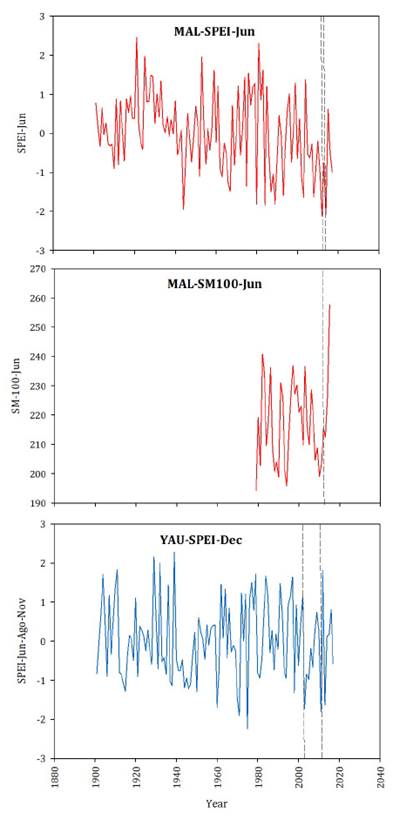
Vertical
lines refer to local extreme drought events.
Las
líneas verticales indican los eventos locales de sequía extrema.
Figure
4.
Historical series of SPEI and SM100 at each site based on correlation function
results.
Figura
4. Series históricas de SPEI y SM100 por cada sitio, basadas
en los resultados de las funciones de correlación.
LFR analysis revealed contrasting responses among stands (figure 5). At YAU, only 6% of trees displayed full resilience to
the 2011 extreme drought. At MAL, no tree exhibited full resilience to the 2010
SM100 event. In contrast, LFR values were comparable between the MAL-SPEI 2012
(LFR = 40%) and the MAL-SPEI 2014 (LFR = 50%) events (figure 5).
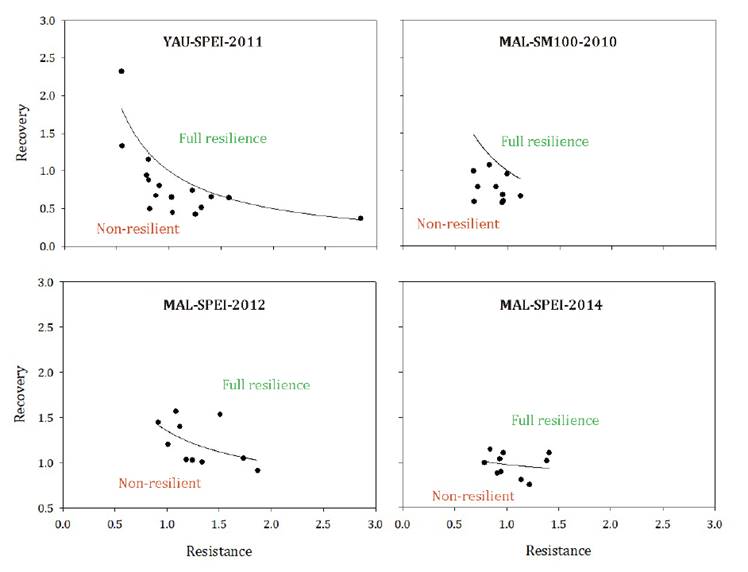
Dots
located above the LFR correspond to trees exhibiting full resilience
(Resilience = 1.0).
Los
puntos situados por encima de la LFR corresponden a los árboles que exhiben
resiliencia plena (Resiliencia = 1.0).
Figure
5.
Comparison of the relationship between observed tree-level mean values of Rt and Rc and the
hypothetical function (black curve) representing the LFR across all Rt values,
for the analyzed extreme atmospheric and edaphic drought events.
Figura
5. Comparación de la relación entre los valores medios
observados a nivel de árbol de Rt y Rc y la función hipotética (curva negra) que
representa la LFR a lo largo de todos los valores de Rt, para los eventos analizados de sequía
atmosférica y edáfica extrema.
Discussion
This study provided novel information of how stone pine growing
in an extra-Mediterranean region responds to atmospheric and edaphic drought
across multiple timescales. Previous research addressed the species’
dendroclimatological signal and resilience to extreme climatic events in Chile
(Loewe-Muñoz et al., 2024ab), but no studies
had explored these dynamics under the environmental conditions of
central-western Argentina. Our findings offer new insights of P. pinea growth
and drought resilience beyond its native Mediterranean habitat, confirming the
species’ adaptability to new climatic contexts.
Although our sample
size was relatively small, it well represented growth dynamics of both
populations. At MAL, sampled trees accounted for 85% of the original plantation
(n = 12). At YAU, the tree-ring chronology reliably captured stand-level
growth, with an Expressed Population Signal (EPS; Wigley
et al., 1984) exceeding the 0.85 threshold, indicating strong
agreement between sampled trees and the overall population (data not shown).
The mean
correlation coefficient among individual series fell within the range reported
in literature, confirming the robustness of our ring-width measurements (Natalini et al., 2016; Piraino
et al., 2013). At YAU, MS was twice as high as at MAL.
Theoretically, MS should reflect MC variability, since both indices are
strongly related (Fritts et al., 1965).
Nevertheless, MC differed only slightly between stands. The higher irrigation
regime at MAL likely explained the differences in MS values by reducing growth
sensitivity to environmental variability. This interpretation agrees with
previous studies in Central European conifer comparing irrigated and
non-irrigated woodlands (Feichtinger et al.,
2014; Rigling et al., 2003).
Mean TRW at both
sites exceeded values previously reported (Mechergui et
al., 2021). This pattern likely reflects both the relatively young age
of the plantations, with trees probably still at their juvenile growth phase,
and the benefit of irrigation, which can extend the growing season, enhance
nutrient mobility, and increase primary production (Feichtinger
et al., 2014; Loewe-Muñoz et al., 2024a). The higher mTRW at
MAL compared with YAU further reflects the more favorable water balance at that
site (precipitation + irrigation: see Materials and Methods).
Our analyses of
drought-radial growth relationships across timescales indicated that irrigation
did not fully decouple growth from climate, suggesting that the supplementary
water was likely insufficient (Perulli et al., 2019).
This limitation was particularly evident at YAU, where radial growth benefited only
during the early phases of ring development. These results have practical
implications for managing future plantations in semi-arid central-west
Argentina. However, the lack of systematic irrigation records prevented
statistical evaluation of watering effect. For future P. pinea plantations
in Mendoza, we recommend monitoring irrigation at least monthly to enable
direct comparison between radial growth dynamics and water supply.
Correlation
function analyses provided quantitative evidence of the species’ distinct
responses to atmospheric and edaphic drought. At MAL, radial growth correlated
with high soil moisture and wet atmospheric conditions at the end of the
growing season. In contrast, at YAU, ring width responded positively to early
summer (December) SPEI. Previous studies in the Mediterranean range reported
similar patterns, showing that radial growth benefits from low
evapotranspiration during the growing season and from abundant precipitation
before cambium reactivation (Mechergui et al.,
2021). Physiologically, P. pinea is a drought-tolerant species,
capable of reducing photosynthetic activity during water stress through root
mortality, stomata control, and biomass allocation (Mechergui
et al., 2021). Nevertheless, drought constrains tree growth by
reducing sap flow and significantly decreasing stem increment (Mechergui et al., 2021; Piraino, 2020). Our
findings confirm that, regardless of irrigation, the species’ radial growth
remains constrained by unfavorable environmental conditions during the growing
period (Loewe-Muñoz et al., 2024b; Piraino, 2020).
Analysis of annual SPEI and SM100 series identified several
extreme drought events that impacted growth dynamics at both sites. At MAL,
edaphic dry spells exerted stronger short-term effects on growth than
atmospheric droughts. This difference may reflect the direct impact of soil
water deficit on roots uptake, whereas atmospheric drought primarily affects
transpiration and leaf water potential, which may not immediately restrict stem
growth if surface soil moisture remains sufficient (Berauer
et al., 2024). Future studies should integrate physiological data
from both root and stem levels, along with site-specific infiltration rates, to
better characterize the species responses to different drought types.
Conclusion
Our study provides critical baseline data on the radial growth
and drought resilience of P. pinea, establishing a foundation for
evaluating its potential for forestry development beyond Mediterranean
environments. These findings are especially relevant for Mendoza Province,
where current forestry focuses almost exclusively on medium-quality wood
production from more water-demanding species like as Populus spp.
Expanding P. pinea cultivation would diversify local forestry and could
yield substantial economic benefits under the semi-arid conditions of
central-western Argentina.
Allen,
C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier,
M., Kitzberger, T., Rigling , A., Breshears, D. D., Hogg, E. H., Gonzalez, P.,
Fensham, R., Zhang, Z., Castro, J., Demidova, N., Lim, J. N., Allard, G., S.,
Running S. W., Semerci, A., Cobb, N. (2010). A global overview of drought and
heat-induced tree mortality reveals emerging climate change risks for forests. Forest
ecology and management, 259(4), 660-684.
Bagnouls,
F. (1953). Saison sèche et indice xérothermique. Bull Soc His nat Toulouse,
88, 193-239.
Barnes,
B. V., Zak, D. R., Denton, S. R., & Spurr, S. H. (1997). IForest ecology
(N° Ed. 4, p. xviii+-774).
Berauer,
B. J., Steppuhn, A., & Schweiger, A. H. (2024). The multidimensionality of
plant drought stress: The relative importance of edaphic and atmospheric
drought. Plant, Cell & Environment, 47(9), 3528-3540.
Biondi,
F., & Waikul, K. (2004). DENDROCLIM2002: A C++ program for statistical
calibration of climate signals in tree-ring chronologies. Computers &
geosciences, 30(3), 303-311.
Calama,
R., Gordo, J., Mutke, S., Conde, M., Madrigal, G., Garriga, E., Arias, M. J.,
Piqué, M., Gandía, R., Monterno, G., Pardos, M. (2020). Decline in commercial
pine nut and kernel yield in Mediterranean stone pine (Pinus pinea L.)
in Spain. Forest-Biogeosciences and Forestry, 13(4), 251.
Calderón,
A., Bustamante, J. A., Riu, N., Perez, S. (2008). Conifers behaviour under
irrigation in the Yaucha dam. Mendoza, Argentina. Revista de la Facultad de
Ciencias Agrarias. Universidad Nacional de Cuyo. 40(1): 67-72. https://bdigital.uncu.edu.ar/app/navegador/?idobjeto=2676
Camarero,
J. J., Gazol, A., Linares, J. C., Fajardo, A., Colangelo, M., Valeriano, C.,
Sánchez-Salguero, R., Sangüesa-Barreda, G., Granda, E., Gimeno, T. E. (2021).
Differences in temperature sensitivity and drought recovery between natural
stands and plantations of conifers are species-specific. Science of the
Total Environment, 796, 148930.
Cook,
E. R., & Krusic, P. J. (2008). A tree-ring standardization program based
on detrending and autoregressive time series modeling, with interactive
graphics (ARSTAN). http://www. ldeo. columbia. edu/res/fac/trl/public/publicSoftware.
html
Diaz
Dentoni, L. (2024). Análisis de la dinámica de crecimiento de Pinus pinea l.
en dos parcelas bajo riego en la provincia de Mendoza. Tesis de grado.
FCA-UNCuyo. 34p.
Di
Rienzo, J., Casanoves, F., Balzarini, M., Gonzalez, L., Tablada, M., Robledo,
C. (2021). InfoStat version 2021.
FAO
(Food and Agriculture Organization). (2020). Global Forest Resources
Assessment 2020: Main report. FAO, Rome.
Feichtinger,
L. M., Eilmann, B., Buchmann, N., & Rigling, A. (2014). Growth adjustments
of conifers to drought and to century-long irrigation. Forest Ecology and
Management, 334, 96-105.
Fritts,
H. C., Smith, D. G., Cardis, J. W., & Budelsky, C. A. (1965). Tree‐ring characteristics along a vegetation
gradient in northern Arizona. Ecology, 46(4), 393-401.
Holmes,
R. L. (1983). Computer-assisted quality control in tree-ring dating and
measurement. Tree-Ring Bulletin, Vol. 43. 1-10.
IPCC
(2023). Sections. In: Climate Change 2023: Synthesis Report. Contribution of
Working Groups I, II and III to the Sixth Assessment Report of the
Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J.
Romero (eds.)]. IPCC. p. 35-115.
Johnstone,
J. F., Allen, C. D., Franklin, J. F., Frelich, L. E., Harvey, B. J., Higuera,
P. E., Mack, M. C., Meentemeyer, R. K., Metz, M. R., Perry, G. L. W.,
Schoennagel, T., Turner, M. G. (2016). Changing disturbance regimes, ecological
memory, and forest resilience. Frontiers in Ecology and the Environment, 14(7),
369-378.
Knutzen,
F., Dulamsuren, C., Meier, I. C., & Leuschner, C. (2017). Recent climate
warming-related growth decline impairs European beech in the center of its
distribution range. Ecosystems, 20, 1494- 1511.
Lloret,
F., Keeling, E. G., & Sala, A. (2011). Components of tree resilience:
effects of successive low‐growth
episodes in old ponderosa pine forests. Oikos, 120(12), 1909-1920.
Loewe-Muñoz,
V., Del Río, R., Delard, C., Balzarini, M. (2022). A detailed time series of
hourly circumference variations in Pinus pinea L. in Chile. Annals of
Forest Science. 79(1): 4.
Loewe-Muñoz,
V., Del Río, R., Delard, C., Cachinero-Vivar, A. M., Camarero, J. J., Navarro-Cerrillo,
R., & Balzarini, M. (2024a). Resilience of Pinus pinea L. trees to
drought in Central Chile based on tree radial growth methods. Forests, 15(10),
1775.
Loewe-Muñoz,
V., Cachinero-Vivar, A. M., Camarero, J. J., Río, R. D., Delard, C., & Navarro-Cerrillo,
R. M. (2024b). Dendrochronological analysis of Pinus pinea in Central Chile and
South Spain for sustainable forest management. Biology, 13(8), 628.
Maxwell,
R. S., & Larsson, L. A. (2021). Measuring tree-ring widths using the
CooRecorder software application. Dendrochronologia, 67, 125841.
Mechergui,
K., Saleh Altamimi, A., Jaouadi, W., & Naghmouchi, S. (2021). Climate change impacts on spatial
distribution, tree-ring growth, and water use of stone pine (Pinus pinea
L.) forests in the Mediterranean region and silvicultural practices to limit
those impacts. iForest- Biogeosciences and Forestry, 14(2),
104.
Mishra,
A. K., & Singh, V. P. (2010). A
review of drought concepts. Journal
of hydrology, 391(1-2), 202- 216.
Muñoz,
V. F. L., Delard, C., González, M. V. G., Mutke, S., & Díaz, V. C. F.
(2012). Introducción del pino piñonero
(Pinus pinea L.) en Chile. Ciencia
& Investigación Forestal, 18(2), 39-52.
Mutke,
S., Calama, R., González-Martínez, S. C., Montero, G., Javier Gordo, F., Bono,
D., & Gil, L. (2012). Mediterranean
stone pine: botany and horticulture. Horticultural
reviews, 39(1), 153-201.
Natalini,
F., Alejano, R., Vázquez-Piqué, J., Pardos, M., Calama, R., & Büntgen, U.
(2016). Spatiotemporal variability of
stone pine (Pinus pinea L.) growth response to climate across the
Iberian Peninsula. Dendrochronologia,
40, 72-84.
Natalini,
F., Alejano, R., Pardos, M., Calama, R., & Vázquez-Piqué, J. (2024). Declining trends in long-term Pinus pinea
L. growth forecasts in Southwestern Spain. Dendrochronologia, 88, 126252.
Navarro-Cerrillo,
R. M., Cachinero-Vivar, A. M., Pérez-Priego, Ó., Cantón, R. A., Begueria, S.,
& Camarero, J. J. (2023). Developing alternatives to adaptive silviculture:
Thinning and tree growth resistance to drought in a Pinus species on an
elevated gradient in Southern Spain. Forest Ecology and Management, 537,
120936.
Perulli,
G. D., Peters, R. L., von Arx, G., Grappadelli, L. C., Manfrini, L., &
Cherubini, P. (2019). Learning from the past to improve in the future:
tree-ring wood anatomy as retrospective tool to help orchard irrigation
management. In IX International Symposium on Irrigation of Horticultural
Crops 1335 (p. 179-188).
Piraino,
S. (2020). Assessing Pinus pinea L. resilience to three consecutive
droughts in central-western Italian Peninsula. iForest-Biogeosciences
and Forestry, 13(3), 246.
Piraino,
S., Camiz, S., Di Filippo, A., Piovesan, G., & Spada, F. (2013). A
dendrochronological analysis of Pinus pinea L. on the Italian
mid-Tyrrhenian coast. Geochronometria, 40, 77-89.
Piraino,
S., Cignoli, F. N., Robledo, S., & Pérez, S. (2021). Un pino mediterráneo
en Sudamérica: situación actual y potencialidad de Pinus pinea L. en
territorio argentino. Experticia, 12(1). 7-10. https://revistas.uncu.edu.ar/ojs3/index.php/experticia/article/view/5041
Piraino,
S., Molina, J. A., Hadad, M. A., & Roig, F. A. (2022). Resilience capacity
of Araucaria araucana to extreme drought events. Dendrochronologia,
75, 125996.
Piraino,
S., Hadad, M. A., Ribas‑Fernández,
Y. A., & Roig, F. A. (2024). Sex-dependent resilience to extreme drought
events: implications for climate change adaptation of a South American
endangered tree species. Ecological Processes, 13(1), 24.
Rigling,
A., Brühlhart, H., Bräker, O. U., Forster, T., & Schweingruber, F. H.
(2003). Effects of irrigation on diameter growth and vertical resin duct
production in Pinus sylvestris L. on dry sites in the central Alps,
Switzerland. Forest Ecology and Management, 175(1-3), 285-296.
Schulman,
E. (1956). Dendroclimatic changes in semiarid America. University of
Arizona.
Schwarz,
J., Skiadaresis, G., Kohler, M., Kunz, J., Schnabel, F., Vitali, V., &
Bauhus, J. (2020). Quantifying growth responses of trees to drought-A critique
of commonly used resilience indices and recommendations for future studies. Current
Forestry Reports, 6, 185-200.
Speer,
J. H. (2010). Fundamentals of tree-ring research. University of Arizona
Press.
Trouet,
V., & Van Oldenborgh, G. J. (2013). KNMI Climate Explorer: a
web-based research tool for high-resolution paleoclimatology. Tree-Ring
Research, 69(1), 3-13.
Vicente-Serrano,
S. M., Beguería, S., & López-Moreno, J. I. (2010). A multiscalar drought
index sensitive to global warming: the standardized precipitation
evapotranspiration index. Journal of climate, 23(7), 1696-1718.
Villalba,
R., & Veblen, T. T. (1997). Spatial and temporal variation in Austrocedrus
growth along the forest steppe ecotone in northern Patagonia. Canadian
Journal of Forest Research, 27(4), 580-597.
Wigley,
T. M., Briffa, K. R., & Jones, P. D. (1984). On the average value of
correlated time series, with applications in dendroclimatology and
hydrometeorology. Journal of Applied Meteorology and climatology, 23(2),
201-213.