Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. En prensa. ISSN (en línea) 1853-8665.
Original article
Statistical Optimization of Seed Coating in Arid-Zone Grass Species Used as Cover Crops in Woody Agroecosystems
Optimización estadística del recubrimiento de semillas en gramíneas de zonas áridas para cultivos de cobertura en agroecosistemas leñosos
Fabio Vazquez 2,
Carlos Alberto Parera 1
1 Universidad Nacional de San Juan (UNSJ). Facultad de Ciencias Exactas, Físicas y Naturales (FCEFyN). Departamento de Biología. Gabinete de Recursos Vegetales. Av. Ig. de la Roza oeste 590. Rivadavia. San Juan. Argentina.
2 Universidad Nacional de San Juan (UNSJ). Facultad de Ingeniería (FI). Av. Libertador Oeste 1109. San Juan. Argentina.
3 Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Godoy Cruz 2290. CABA. Argentina.
* caromeglioli@gmail.com
Abstract
Inter-row vegetative cover improves soil physicochemical properties and biodiversity but should not compete with main crop for water. Native grass species such as Leptochloa crinita (Lag.) P. M. Peterson & N. W. Snow, Pappophorum caespitosum R. E. Fr. and Digitaria californica (Benth.) Henrard var. californica provide forage value, require less water than conventional cover crops, and exhibit high tolerance to the edaphoclimatic conditions of arid regions. However, their small seed size hinders efficient handling and sowing. Seed coating increases seed size and weight, forming a pellet that improves handling, distribution, and protection. Nevertheless, limited information is available on coating techniques for small-seeded species. This study aims to optimize the coating process through a statistically designed experiment to evaluate key morphological and physiological seed traits. A Box-Behnken design was applied to optimize one compositional factor of the pellet and two operational parameters. Pellet traits such as size, heterogeneity, seed number, and residue were analyzed alongside germination and seedling development. Optimization was first carried out using edible amaranth seeds as a model organism and then validated with native grasses. The resulting models were statistically significant and allowed the identification of an optimal operational range. Germination was unaffected in single-seed pellets, and root growth was enhanced by the coating process. Overall, the proposed statistical methodology proved effective and broadly applicable for optimizing small-seed coating processes.
Keywords: Native seeds, Leptochloa crinita, Pappophorum caespitosum, Digitaria californica
Resumen
Las coberturas vegetales interfilares mejoran las propiedades fisicoquímicas y la biodiversidad del suelo, pero no deberían competir por el agua en los cultivos. Algunas especies nativas, como Leptochloa crinita (Lag.) P. M. Peterson & N. W. Snow, Pappophorum caespitosum R. E. Fr. y Digitaria californica (Benth.) Henrard var. californica, ofrecen valor forrajero, requieren menos agua que cultivos de cobertura convencionales y presentan alta tolerancia a las condiciones edafoclimáticas del árido. No obstante, el pequeño tamaño de sus semillas dificulta su manejo. El recubrimiento incrementa tamaño y peso de las mismas, formando un pellet que facilita su manipulación, distribución y protección. Sin embargo, hay escasa información sobre el recubrimiento de semillas pequeñas. Este estudio busca optimizar dicho proceso mediante diseño experimental y evaluar variables morfológicas y fisiológicas clave. Se aplicó un diseño Box-Behnken para optimizar un factor de composición del pellet y dos factores operativos. Se evaluaron características del pellet como tamaño, heterogeneidad, cantidad de semillas y residuo, así como la germinación y desarrollo de plántulas. La optimización se realizó con semillas de amaranto de consumo humano como organismo modelo y fue validada en gramíneas nativas. Los modelos resultaron significativos y permitieron definir una zona operativa óptima. La germinación no se vio afectada en pellets de una semilla, más aún el crecimiento radicular se vio beneficiado por el proceso de recubrimiento. Finalmente, la metodología estadística utilizada demostró ser aplicable y eficaz para optimizar el recubrimiento de semillas.
Palabras clave: Semillas nativas, Leptochloa crinita, Pappophorum caespitosum, Digitaria californica
Originales: Recepción: 26/06/2025- Aceptación: 09/12/2025
Introduction
Some agronomic management practices in woody crops like vineyards, olive groves, almond orchards, and walnut plantations can act as regionally significant drivers of soil degradation. Inter-row areas in these systems, particularly under pressurized irrigation, are often classified as “degraded areas” due to prolonged soil exposure to erosive forces (Chen et al. 2010).
Inter-row cover crops provide multiple agro-environmental benefits reducing weed competition, enhancing biodiversity, improving nutrient cycling, and serving as green manure (Rubio-Asensio et al. 2022; Thioye et al. 2021; Tomaz et al. 2021). However, in semi-arid regions, conventional cover crops may compete with main crop for water and nutrients. This limitation encourages the use of native species, which generally possess deep root systems, lower water requirements, and superior adaptation to local edaphoclimatic conditions (Pornaro et al. 2022; Massa Decon & Pérez, 2025).
Native grasses such as Leptochloa crinita (Lag.) P. M. Peterson & N. W. Snow, Pappophorum caespitosum R. E. Fr. and Digitaria californica (Benth.) Henrard var. californica are valued for their tolerance to drought, aridity and salinity, rapid establishment, and low water use (Kozub et al. 2018; Vega-Riveros et al. 2020). D. californica has already been used successfully as an inter-row species (Ferrari & Parera, 2015), while L. crinita and P. caespitosum have been employed as forage grasses (Marinoni et al. 2019). Nevertheless, these native species produce small, lightweight seeds (often less than 1 mm), with variable shapes (Carballo et al. 2005; Ramírez-Segura et al. 2022), making them difficult to handle and sow (Pedrini et al. 2017). Germination rates also improve significantly when floral residues are removed, exposing caryopsis (Meglioli et al. 2024).
Seed coating intends to overcome these challenges. By enclosing the caryopsis within a protective layer, coating increases seed size, enhances mechanical and distribution, and reduces damage during sowing (Afzal et al. 2020; Pedrini et al. 2017). Moreover, it can improve germination and plant establishment (Pedrini et al. 2017). Coating mixtures of native species may also improve agroecosystem resilience and stability by increasing functional diversity (Madsen et al. 2012).
Although seed coating technologies have been primarily developed for major agricultural species (Pedrini et al. 2017), there is limited literature regarding suitable coating material and operational parameters for small native grasses. In addition, many studies on seed coating lack robust experimental design (Rajeshwari et al. 2020). Statistical experimental design and response surface methodology provide powerful tools to analyze factor interactions, reduce experimental effort, and model complex biological responses (Pedrozo et al. 2024; Leardi, 2009).
Here, we hypothesized that the coating process would not negatively affect the germination of L. crinita, P. caespitosum, and D. californica. To test this hypothesis, the present study optimized key factors in the mechanical coating of small amaranth seeds a model organism due to its similar morphology (Zubillaga et al. 2024). The optimized conditions were then validated with the target native grasses by evaluating pellet size, seed conglomeration, and germination performance.
This study provides an original integrative approach, combining statistical optimization with the use of a model organism. Focusing on seed-level responses, the research explores variability and highlights how pellet composition and seed number influence germination and early seedling development.
Materials and Methods
Plant Material
Because obtaining spikelet-free grass seeds is a labor-intensive process and a consistent seed supply was required for process optimization, commercial amaranth seeds were used as a model due to their morphological similarity to native grasses (Xavier et al. 2019).
For validation, seeds of L. crinita, P. caespitosum, and D. californica (Poaceae) were provided by the native species germplasm bank (ARG1416) at the Plant Resources Cabinet from Universidad Nacional de San Juan (UNSJ). Seeds were mechanically scarified and air-cleaned to obtain naked caryopses.
Seed Coating Optimization Procedure and Factors
Lavanya et al. (2011) described for mechanisms of pellet formation: nucleation, coalescence, layering, and abrasion. In the present study, nucleation was excluded because the seed served as the nucleus. Coalescence refers to the formation of larger particles from multiple nuclei; layering involves the successive addition of powder layers; and abrasion encompasses the material exchange between forming particles.
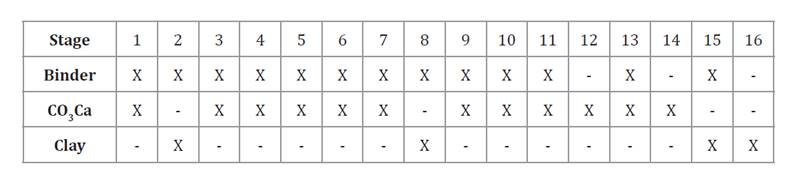
Based on previous reports and acknowledging that the sequential addition of filler and binder significantly affects final pellet size (Rajeshwari et al. 2020), the process was structured into 16 coating stages (table 1).
Table 1. Seed coating stages.
Tabla 1. Etapas del recubrimiento de semillas.
Quantitative and qualitative properties of the coating materials (table 2) were selected following ecological and sustainability criteria, key principles for ecological restoration (Hoose et al. 2019).
Table 2. Quantitative and qualitative factors considered.
Tabla 2. Factores cuantitativos y cualitativos considerados.
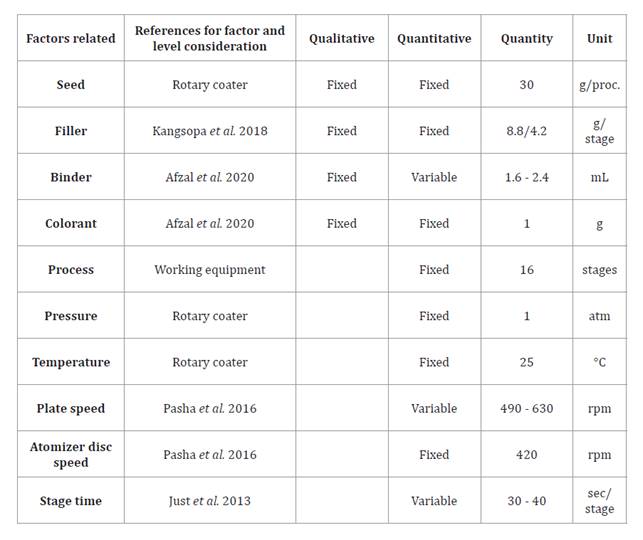
* Factors were considered under ecological restrictions
* Los factores se evaluaron dentro de restricciones ecológicas.
Filler: A 70:30 (w/w) mixture of calcium carbonate and diatomaceous earth (Kangsopa et al. 2018). White clay (kaolin) was used as a surface-smoothing agent (Romo-Campos & Guzmán Valle, 2019). Binder: Gum arabic (25% w/v), a natural water-soluble polysaccharide with a viscosity of 156.7 cP at 25°C, was used. Colorant: A food-grade powder dye was added for visual differentiation. After reviewing literature, it was decided to optimize three independent variables: plate rotational speed (490, 560, and 630 rpm), binder volume (1.6, 2, and 2.4 mL), and stage time (30, 35, and 40 s).
Evaluated Responses on the Optimization and Validation
Thirty pellets were randomly selected for measurement. Pellet size was determined by recording the major and minor diameters (mm) using a digital caliper. Size heterogeneity was calculated as the standard deviation of these diameters. To determine the number of seeds per pellet, each pellet was gently crushed, and the enclosed seeds were counted under a stereomicroscope. The amount of residue was quantified by collecting and weighing (g) the material adhered to the drum and plate of the rotary coater after coating. This value was subtracted from the total input weight of filler and binder to estimate the proportion of material loss.
Numerical Optimization of Multiple Responses
Using the fitted models, a numerical multi-response optimization was performed. The objectives were to: maximize pellet size, minimize the number of seeds per pellet (target = 1), minimize size heterogeneity, and minimize residue amount.
Pellet Validation
A single optimal point derived from the numerical optimization was selected to validate the desirability function and predictive models. For this purpose, a mixture of 30 g of cleaned caryopses was prepared, comprising 10 g each of L. crinita, P. caespitosum, and D. californica.
Additional Responses Evaluated during Validation
Dissolution: Pellets were placed on germination paper moistened and saturated with distilled water. The time required for complete disintegration was recorded.
Fragmentation: Pellets were placed in a 15 mL Falcon tube and subjected to continuous vibration for 1 min using a vortex shaker (Arcano) at 800 rpm. The number of broken pellets was counted (modified from Rajeshwari et al. 2020).
Germination Responses
To evaluate the effect of coating on germination, 30 pellets were sown in sterile Petri dishes lined with germination paper moistened with 6 mL of distilled water. Treatments included (four replicates each): caryopses (control), consisting of 10 seeds from each native species; single-seed pellets, diameter < 1.6 mm; and multi-seed pellets (conglomerates), diameter between 1.6 and 2.8 mm, containing an average of five seeds per pellet (150 seeds total) (Hoose et al. 2019). Plates were incubated in a germination chamber at 25°C under a 12 h light/dark cycle for five days. Germination was recorded when radicle emergence was visible, and shoot and radicle lengths were measured with a caliper.
Equipment
The coating process was performed using a rotary coater (SATEC Concept model ML 2000, SATEC Equipment GmbH, Germany (figure 1).
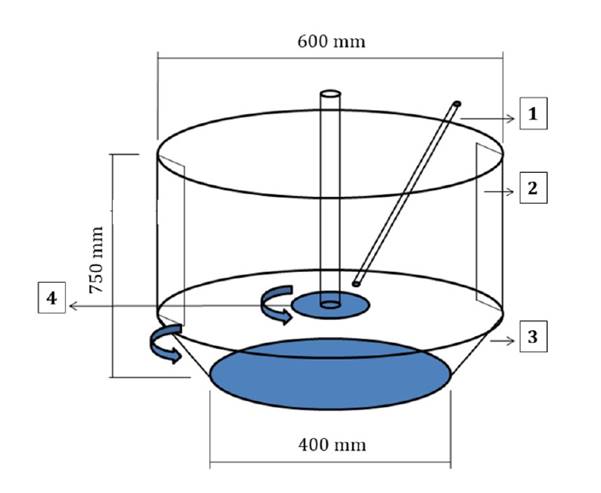
1- liquid pipeline, 2- baffles, 3- rotary coater plate, 4- atomizer disc.
1- Conducto para líquido. 2- Deflectores. 3- Plato giratorio recubridor. 4- Disco atomizador.
Figure 1. Schematic representation of the rotary coater (SATEC Concept model ML 2000).
Figura 1. Esquema interno. Modelo Concept SATEC ML 2000.
Statistical Analysis
All experimental design (Box-Behnken) and analyses were performed using Design-Expert 7.0.0 (Stat-Ease Inc., USA). Model significance, lack of fit, adequacy, precision, robustness, and numerical optimization were assessed. ANOVA assumptions, outliers, and data transformations were also analyzed to ensure model validity.
Results
Seed Coating Optimization
Table 3 summarizes the factor level used in the Box–Behnken experimental design and the corresponding responses obtained from the coating process for each experimental run.
Table 3. Box–Behnken design showing variation in plate speed, stage duration, and binder volume, and their corresponding responses: pellet size, seed number, residue amount, and size heterogeneity.
Tabla 3. Diseño Box-Behnken: Variación de factores: velocidad del plato, tiempo por etapa y cantidad de aglutinante; y resultados de la optimización: tamaño del pellet, cantidad de semillas, cantidad de residuo y heterogeneidad del tamaño.
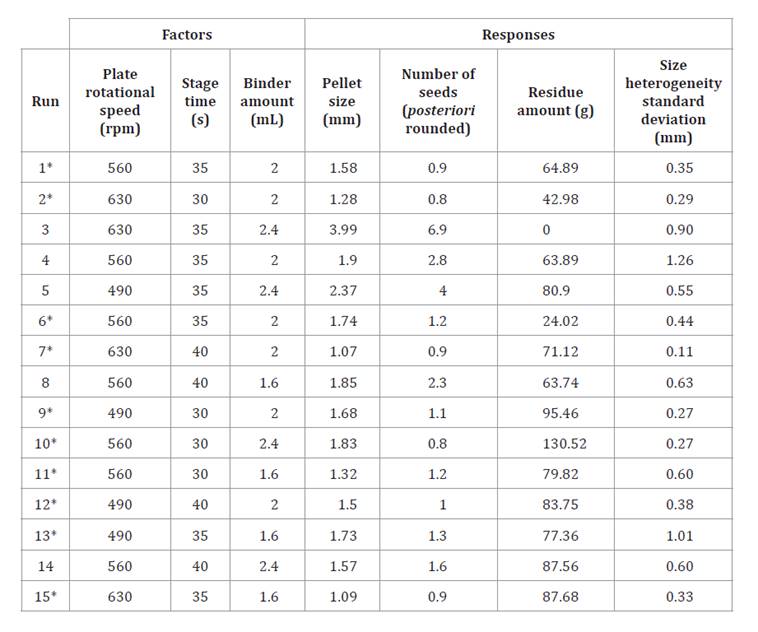
* Runs for single-seed pellet size model.
* Corridas para el modelo de tamaño de pellets de una semilla.
Modeling of Responses
Pellet size data were analyzed in two subsets: (A) single-seed pellets (figure 2A) and (B) all data combined (figure 2B), including multi-seed (conglomerate) pellets (Hoose et al. 2019).
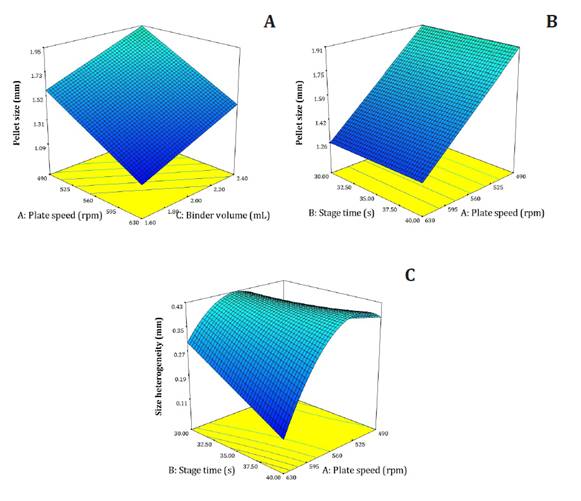
(A) Single-seed pellets; (B) combined dataset (single- and multi-seed pellets); (C) size heterogeneity (standard deviation) of single-seed pellets. Binder volume fixed at 2 mL.
A- Tamaño del pellet con una sola semilla. B- Todos los datos, tamaños de pellets con una y múltiples semillas. C- Heterogeneidad del tamaño en pellets de una sola semilla (desviación estándar). Volumen de aglutinante fijo en 2 mL.
Figure 2. Influence of the evaluated factors on pellet size.
Figura 2. Influencia de los factores evaluados sobre el tamaño del pellet.
All response models were significant (table 4) and showed no lack of fit unless otherwise indicated. Adjusted and predicted R² values were consistent, and ANOVA assumptions were met.
Table 4. Responses and models/ Statistical summary of the fitted models.
Tabla 4. Respuestas y modelos/ Resumen estadístico de los modelos ajustados.
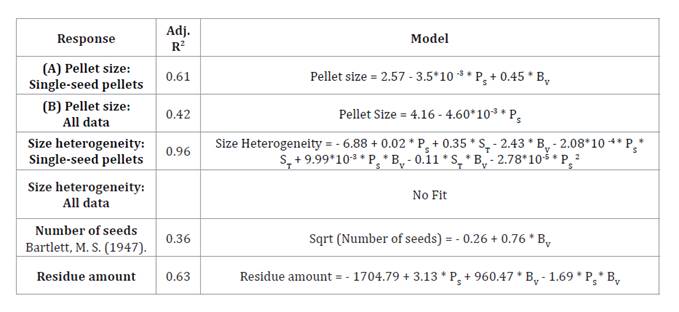
* (PS: Plate speed, BV: Binder volume, ST: Stage time)
* (PS: Velocidad del plato, BV: Volumen del aglutinante, ST: Tiempo de etapa).
Pellet Size and Size Heterogeneity
Models indicated that increasing the rotational speed of the rotary coater plate reduced pellet size (figure 2A-B). Conversely, greater binder volume increased pellet size for single-seed pellets (figure 2A).
Size heterogeneity (figure 2C) was analyzed only for single-seed pellets, since combining single-and multi-seed data was better represented by the overall mean (mean = 0.54). Longer stage duration and higher rotation speed reduced size variability among pellets.
Number of Seeds per Pellet
The model describing the number of seeds per pellet was significant (adjusted R² = 0.36; figure 3). Although predictive precision was moderate, the model adequately represented overall behavior (Pedrozo et al. 2024). Increasing binder volume led to the formation of conglomerates.
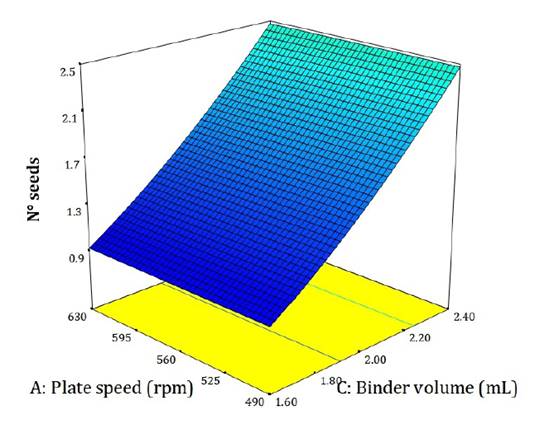
Figure 3. Influence of the evaluated factors on the number of seeds per pellet.
Figura 3. Influencia de los factores evaluados sobre la respuesta de cantidad de semillas.
Amount of Residue
During coating, some material adhered to the rotary drum and plate, contributing to residue accumulation (figure 4).
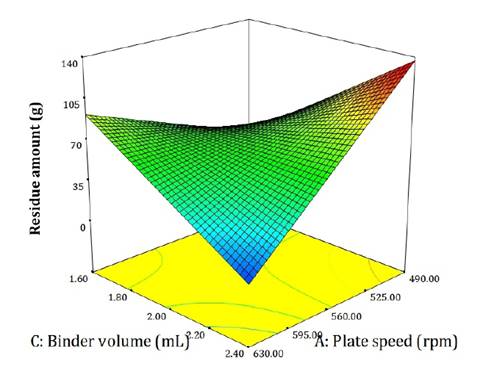
Figure 4. Effect of evaluated factors on the residue adhered to the rotary coater drum and plate at the end of process.
Figura 4. Influencia de los factores evaluados sobre la cantidad de residuo adherido al tambor y al plato de la peletizadora al final del proceso.
The smallest amounts of residue occurred at high plate rotation speed and maximum binder volumes, which coincided with the formation of conglomerates. Conversely, low plate speeds combined with high binder volumes produced the largest residues.
Residue per experiment ranged from 0 to 140 g, averaging 43% of total input mass.
Multi-Response Optimization
A simultaneous optimization was performed using desirability criteria for the following responses: pellet size (maximize, importance: +++), size heterogeneity (minimize, +), number of seeds (minimize, ++++), and amount of residue (minimize, ++). The optimal compromise solution is shown in figure 5. All models used corresponded to the complete dataset (single- and multi-seed pellets).
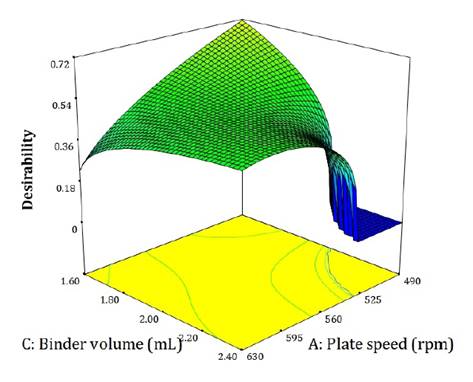
Pellet size (maximize +++), size heterogeneity (minimize+), amount of seed (minimize +++++), and amount of residue (minimize ++).
Tamaño del pellet (maximizar +++), Heterogeneidad del tamaño (minimizar +), Cantidad de semillas (minimizar +++++), y Cantidad de residuo (minimizar ++).
Figure 5. Multiple-response optimization based on experimenter-defined criteria.
Figura 5. Optimización de múltiples respuestas según criterios del experimentador.
Maximum desirability was achieved at low plate speed and low binder volume. A marked decrease in desirability occurred under conditions of low plate speed and high binder volumes, corresponding to zones of formation of conglomerates and high residue accumulation.
Prediction and Validation
Model predictions were validated using the following optimized conditions: plate speed = 595 rpm, binder volume = 2.2 mL, and stage duration = 35 s (overall desirability: 0.51). Validation used caryopses of L. crinita, P. caespitosum, and D. californica (table 5).
Table 5. Validation responses: Pellet size, size heterogeneity, and number of seeds after classification into four size categories, expressed as weight percentage of the total mix. Additional responses: Fragmentation and dissolution time.
Tabla 5. Respuestas de validación: Tamaño del pellet, heterogeneidad del tamaño y número de semillas tras el tamizado en cuatro tamaños, y porcentaje de representación del total de la mezcla en peso. Respuestas adicionales: Fragmentación y disolución.
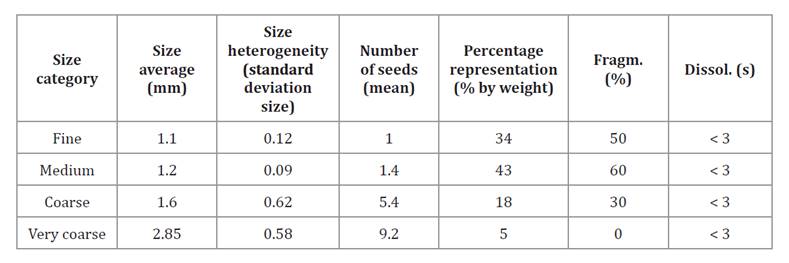
* Fragm.: violent fragmentation; Dissol.: pellet dissolution time.
* Fragm.: Fragmentación violenta, Dissol.: Tiempo de disolución del pellet.
Pellets were classified into four size categories, and validation results were calculated as weighted means based on proportional mass. Two additional responses were also evaluated: fragmentation and dissolution time.
Weighted means: pellet size = 1.38 mm; number of seeds per pellet = 3.15; heterogeneity (SD) = 0.36. Single-response value: amount of residue = 25% at the end of the process.
Predicted values for these conditions are presented in table 6.
Table 6. Model-predicted responses under optimal conditions (plate speed 595 rpm; binder volume 2.2 mL; stage time 35 s).
Tabla 6. Predicción de las respuestas del pellet bajo las condiciones: velocidad del plato (595 rpm), volumen de aglutinante (2,2 mL), tiempo por etapa (35 segundos).
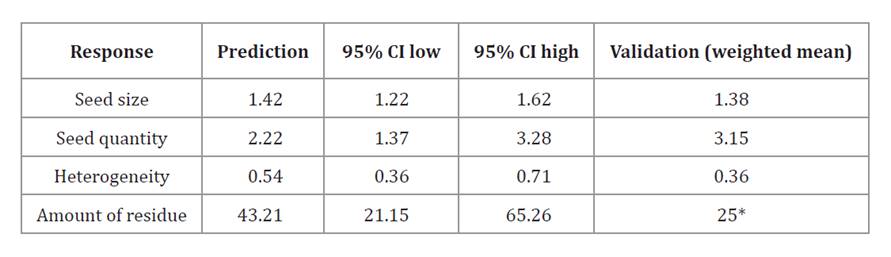
* Single response, not weighted mean.
* Respuesta individual, no promedio ponderado.
All validation results fell within the 95% confidence intervals predicted by the models.
Germination and Seedling Performance
Pellets obtained during validation (table 6) were used to evaluate germination and early seedling development in native grasses (L. crinita, P. caespitosum, and D. californica).
Germination
No significant differences were detected in germination percentage between control caryopses and single-seed pellets (figure 6). However, multi-seed (conglomerate) pellets showed a 20% reduction compared to both control and single-seed treatments.
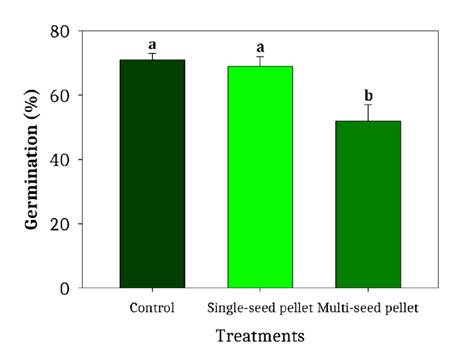
Bars indicate: standard deviation; different letters denote significant differences (Fisher’s LSD, α ˂ 0.05).
Las barras indican la desviación estándar. Letras diferentes indican diferencias significativas (LSD de Fisher, α ˂ 0,05).
Figure 6. Germination percentages for caryopses (control), single-seed pellets, and multi-seed pellets. Species: L. crinita, P. caespitosum, and D. californica.
Figura 6. Porcentajes de germinación obtenidos en los tratamientos de cariopses (control), pellets de una semilla y pellets de múltiples semillas. Especies: L. crinita, P. caespitosum y D. californica.
Shoot and Radicle Length
Shoot and radicle length results are shown in figure 7. Shoot length did not differ significantly among treatments. In contrast, radicle length was significantly greater in pelleted seeds (2.12 ± 0.17 cm) than in controls (1.78 ± 0.11 cm).
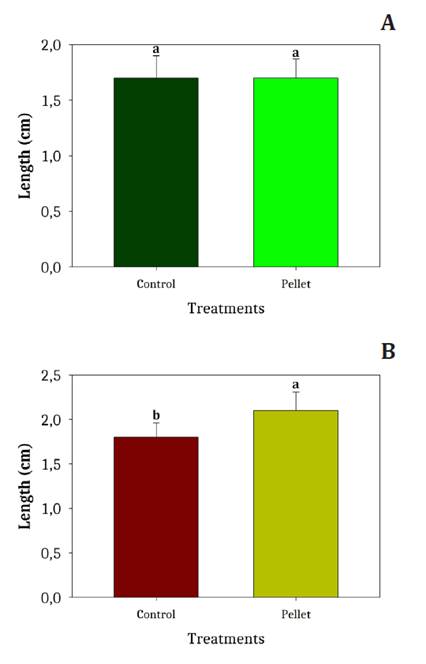
Bars represent standard deviation; different lowercase letters indicate significant differences (Fisher’s LSD, α ˂ 0.05).
Las barras indican la desviación estándar. Letras minúsculas diferentes indican diferencias significativas (LSD de Fisher, α ˂ 0,05).
Figure 7. (A) Shoot and (B) radicle length in caryopses (control) and pellet treatments for L. crinita, P. caespitosum, and D. californica.
Figura 7. (A) Longitud de plántula y (B) longitud de radícula para los tratamientos: cariopses (control) y pellets. Especies: L. crinita, P. caespitosum y D. californica.
Discussion
This study provides one of the first comprehensive descriptions of the seed-coating process for small-seeded species, detailing component quantities, proportions, processing times, and operational stages. It also represents the first statistical optimization of seed coating for native grass from arid regions. Although seed-coating technologies have been developed for decades, detailed methodological information is rarely available in the public domain, as most advances are confined to private industry (Pedrini et al. 2017).
Optimization of Individual Responses
The fitted models showed that the rotational speed of the coater plate had a significant influence on pellet size and homogeneity. The shear effect associated with higher speeds likely caused a reduction in pellet size (figures 2A-B) due to material detachment from the forming pellet (Iparraguirre, 2009; Pasha et al. 2016). Conversely, increasing binder volume in single-seed pellets resulted in larger pellet (figure 2A), in agreement with reports from the pharmaceutical industry where higher impeller speeds reduce pellet size, whereas binder concentration increases it (Thies & Kleinebudde, 1999). Longer stage durations were associated with reduced size heterogeneity (figure 2C), in line with the findings of Just et al. (2013). The combination of layering and abrasion appears to benefit from extended processing times, promoting more uniform pellets. Heterogeneity also decreased with increasing plate speed, confirming the observations made by Pasha et al. (2016). Regarding the number of seeds per pellet (figure 3), higher binder viscosity induced by increased binder volume promoted seed adhesion and coalescence (Johansen & Shaefer, 2001; Keningley et al. 1997; Lavanya et al. 2011). For residue response (figure 4), a relevant factor considering the effort required to obtain spikelet-free native grass seeds (Meglioli et al. 2024), the combination of greater conglomerate mass and higher rotational speed enhanced the detachment of adhering material, reducing residue (Iparraguirre, 2009). In contrast, low speed combined with high binder volume increased adhesion to the plate and drum, resulting in higher residue levels (Johansen & Shaefer, 2001). The mean residue of 43% per run was lower than that reported by Pedrini et al. (2017) for Solanum lycopersicum (65.3%) and Microlaena stipoides (60.9%).
Multi-Response Optimization, Validation, and Germination in Native Seeds
Multi-response desirability analysis successfully identified optimal coating conditions. Validation confirmed both the predictive accuracy and robustness of the statistical models, as all measured values for native grass seeds fell within the 95% confidence intervals of model predictions.
Seed coating can enhance or impair seed physiology by facilitating water and nutrient uptake or, conversely, by impeding radicle emergence (Pedrini et al. 2017; Sprey et al. 2019). In the present study, germination of single-seed pellets was unaffected, in agreement with the results obtained using comparable materials (Romo-Campos & Guzmán Valle, 2019). In contrast, multi-seed pellets (conglomerates) showed reduced germination, possibly due to allelopathic interactions among seeds during germination, as reported in other grass species (Möhler et al. 2018). The enhanced radicle length observed in pelleted seeds may be associated with the coating components (CaCO₃, diatomaceous earth, kaolin), which provide calcium and silicon, elements linked to cell elongation, improved nutrient transport, and stress tolerance (Bauer et al. 2011; Jalal et al. 2023). Additionally, the high intraspecific genetic variability of native grasses (Trod et al. 2018; Villagra et al. 2011), combined with the mixed caryopses from three species, could explain the variability seen in physiological responses.
Conclusions
Validation using small-seeded native grasses confirmed the applicability, accuracy, and robustness of the statistical optimization methodology. All pellet traits fell within the predicted confidence intervals, supporting the reliability of the models. Seedling evaluations indicated that coating did not negatively affect germination, possibly due to the rapid dissolution of the coating material, and that the process positively influenced radicle elongation.
Afzal, I., Javed, T., Amirkhani, M., Taylor, A. G. (2020). Modern Seed Technology: Seed Coating Delivery Systems for Enhancing Seed and Crop Performance. Agriculture, 10, 526. https://doi. org/10.3390/agriculture10110526
Bartlett, M. S. (1947). The Use of Transformations. Biometrics, 3(1): 39-52. https://doi. org/10.2307/3001536
Bauer, P., Elbaum, R., Weiss, I. M. (2011). Calcium and silicon mineralization in land plants: transport, structure and function. Plant Science, 180(6), 746-756. https://doi.org/10.1016/j. plantsci.2011.01.019
Carballo, D. J., Matus, M., Betancourt, M., Ruiz, C. (2005). Manejo de Pasto I. Facultad de Ciencia Animal. Universidad Nacional Agraria. https://repositorio.una.edu.ni/id/ eprint/2425
Chen, D., Suter, H. C., Islam, A., & Edis, R. (2010). Influence of nitrification inhibitors on nitrification and nitrous oxide (N2O) emission from a clay loam soil fertilized with urea. Soil Biology Biochem, 42(4): 660-664. https://doi.org/10.1016/j.soilbio.2009.12.014
Ferrari, F. N., & Parera, C. A. (2015). Germination of six native perennial grasses that can be used as potential soil cover crops in drip-irrigated vineyards in semiarid environs of Argentina. Journal of Arid Environments, 113, 1-5. https://doi.org/10.1016/j.jaridenv.2014.09.002
Hoose, B. W., Call, R. S., Bates, T. H., Anderson, R. M., Roundy, B. A., Madsen, M. D. (2019). Seed conglomeration: a disruptive innovation to address restoration challenges associated with small-seeded species. Restoration Ecology, 27(5): 959-965. https://doi.org/10.1111/ rec.12947
Iparraguirre, L. M. (2009). Mecánica Básica: fuerza y movimiento. Instituto Nacional de Educación Tecnológica. http://www.bnm.me.gov.ar/giga1/ documentos/EL001845.pdf
Jalal, A., da Silva Oliveira, C. E., Galindo, F. S., Rosa, P. A. L., Gato, I. M. B., de Lima, B. H., Teixeira Filho, M. C. M. (2023). Regulatory Mechanisms of Plant Growth-Promoting Rhizobacteria and Plant Nutrition against Abiotic Stresses in Brassicaceae Family. Life, 13, 211. https://doi. org/10.3390/ life13010211
Johansen, A., Schæfer, T. (2001). Effects of interactions between powder particle size and binder viscosity on agglomerate growth mechanisms in a high shear mixer. European Journal of Pharmaceutical Sciences, 12(3), 297-309. https://doi.org/10.1016/S0928- 0987(00)00182-2
Just, S., Toschkoff, G., Funke, A., Djuric, D., Scharrer, G., Khinast, J., Knop, K., Kleinebudde, P. (2013). Optimization of the inter-tablet coating uniformity for an active coating process at lab and pilot scale. International journal of pharmaceutics, 457(1), 1-8. http://dx.doi. org/10.1016/j. ijpharm.2013.09.010
Kangsopa, J., Hynes, R. K., Siri, B. (2018). Lettuce seeds pelleting: A new bilayer matrix for lettuce (Lactuca sativa) seeds. Seed Science and Technology, 46(3), 521-531. https://doi. org/10.15258/ sst.2018.46.3.09
Keningley, S. T., Knight, P. C., Marson, A. D. (1997). An investigation into the effects of binder viscosity on agglomeration behaviour. Powder Technology, 91(2), 95-103. https://doi.org/10.1016/ S0032-5910(96)03230-5
Kozub, P. C., Cavagnaro, J. B., Cavagnaro, P. F. (2018). Exploiting genetic and physiological variation of the native forage grass Trichloris crinita for revegetation in arid and semi‐arid regions: An integrative review. Grass and Forage Science, 73(2), 257-271. https://doi.org/10.1111/ gfs.12337
Lavanya, K., Senthil, V., Rathi, V. (2011). Pelletization technology: a quick review. International Journal of Pharmaceutical Sciences and Research, 2(6), 1337. http://dx.doi.org/10.13040/ IJPSR.0975-8232.2(6).1337-55
Leardi, R. (2009). Experimental design in chemistry: A tutorial. Analytica Chimica Acta, 652(1-2), 161- 172. https://doi.org/10.1016/j.aca.2009.06.015
Madsen, M. D., Davies, K. W., Jason Williams, C. & Svejcar, T. J. (2012). Agglomerating seeds to enhance native seedling emergence and growth. Journal of Applied Ecology, 49, 431-438 https:// doi.org/10.1111/j.1365-2664.2012.02118.x
Marinoni, L. D. R., Zabala, J. M., Taleisnik, E. L., Schrauf, G. E., Richard, G. A., Tomas, P. A., Giavedoni, J. A. & Pensiero, J. F. (2019). Wild halophytic species as forage sources: Key aspects for plant breeding. Grass and Forage Science, 74(3), 321-344. https://doi.org/10.1111/gfs.12410
Massa Decon, A. M.; Pérez, S. 2025. Nursery Production of Neltuma Genus in Arid and Semiarid Regions of Argentine: a Review. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 57(2): 211-223. DOI: https://doi.org/10.48162/rev.39.184
Meglioli, C., Scaglia, J. A., Parera, C. A. (2024). Factores que afectan la germinación de tres pastos de zonas áridas: Leptochloa crinita, Pappophorum caespitosum y Digitaria californica (Poaceae). Acta Botanica Mexicana, (131). https://doi.org/10.21829/abm131.2024.2329
Möhler, H., Diekötter, T., Herrmann, J. D., & Donath, T. W. (2018). Allelopathic vs. autotoxic potential of a grassland weed-evidence from a seed germination experiment. Plant Ecology & Diversity, 11(4), 539–549. https://doi.org/10.1080/17550874.2018.1541487
Pasha, M., Hare, C., Ghadiri, M., Gunadi, A., Piccione, P. M. (2016). Effect of particle shape on flow in discrete element method simulation of a rotary batch seed coater. Powder Technology, 296, 29-36. http://dx.doi.org/10.1016/j.powtec.2015.10.055
Pedrini, S., Merritt, D. J., Stevens, J., Dixon, K. (2017). Seed Coating: Science or Marketing Spin? Trends in Plant Science, 22(2): 106-116. https://doi.org/10.1016/j.tplants.2016.11.002
Pedrozo, L. P., Kuchen, B., Flores, C. B., Rodríguez, L. A., Pesce, V. M., Maturano, Y. P., Nally, M. C., Vazquez, F. (2024). Optimization of sustainable control strategies against blue rot in table grapes under cold storage conditions. Postharvest Biology and Technology, 213, 112946. https:// doi. org/10.1016/j.postharvbio.2024.112946
Pornaro, C., Meggio, F., Tonon, F., Mazzon, L., Sartori, L., Berti, A., Macolino, S. (2022). Selection of inter-row herbaceous covers in a sloping, organic, non-irrigated vineyard. PLoS ONE, 17(12): e0279759. https://doi.org/10.1371/journal.pone.0279759
Rajeshwari, M., Renganayaki, P. R., Vanitha, C., Ravikesavan, R. (2020). Standardization of Pellet size for Mechanized Sowing in Foxtail Millet (Setaria italica). Madras Agricultural Journal. https:// doi.org/10.29321/MAJ.2020.000370
Ramírez-Segura, E., Maldonado-Jaquez, J. A., Torres-Salas, I., Quero-Carrillo, A. R. (2022). Atributos físicos y fisiológicos de semilla de pastos nativos del México árido. Revista fitotecnia mexicana, 45(4): 453-460. https://doi.org/10.35196/rfm.2022.4.453
Romo Campos, R. de L., & Guzmán Valle, J. F. G. (2019). Recubrimiento de semillas como técnica para la restauración de suelos degradados. e-CUCBA 11: 46-53. https://doi.org/10.32870/e-cucba.v0i11.128
Rubio-Asensio, J. S., Abbatantuono, F., Ramírez-Cuesta, J. M., Hortelano, D., Ruíz, J. L, Parra, M., Martínez-Meroño, R. M., Intrigliolo, D. S., Buesa, I. (2022). Effects of cover crops and drip fertigation regime in a young almond agroecosystem. Agronomy, 12(11), 2606. https:// doi.org/10.3390/ agronomy12112606
Sprey, L. M., Ferreira, S. A. D. N., Sprey, M. M. (2019). Physiological quality of pelleted Cubiu (Solanum sessiliflorum Dunal) seeds. Revista Brasileira de Fruticultura, 41, e-075. https://doi. org/10.1590/0100-29452019075
Thies, R., Kleinebudde, P. (1999). Melt pelletisation of a hygroscopic drug in a high shear mixer: Part 1. Influence of process variables. International journal of pharmaceutics, 188(2), 131-143. https://doi.org/10.1016/S0378-5173(99)00214-8
Thioye, B., Legras, M., Castel, L., Hirissou, F., Chaftar, N., Trinsoutrot-Gattin, I. (2021). Understanding arbuscular mycorrhizal colonization in walnut plantations: The contribution of cover crops and soil microbial communities. Agriculture, 12(1), 1. https://doi.org/10.3390/ agriculture12010001
Tomaz, A., Coleto Martínez, J., Arruda Pacheco, C. (2021). Effects of cover crops and irrigation on ‘Tempranillo’ grapevine and berry physiology: an experiment under the Mediterranean conditions of Southern Portugal. OENO One, 55(3), 191-208. https://doi.org/10.20870/ oeno-one.2021.55.3.4629
Trod, M. A., Tomas, P. A., Cervigni, G. D., Zabala, J. M., Marinoni, L. D., Giavedoni, J. A. (2018). Análisis de la distribución y densidad estomática en Trichloris (Chloridoideae, Poaceae), bajo diferentes regímenes hídricos. Boletín de la Sociedad Argentina de Botánica, 53(4): 1-10. http:// dx.doi.org/10.31055/1851.2372.v53.n4.21979
Vega Riveros, C., Villagra, P. E., & Greco, S. A. (2020). Different root strategies of perennial native grasses under two contrasting water availability conditions: implications for their spatial distribution in desert dunes. Plant Ecology, 221(7), 633-646
Villagra, P. E., Giordano, C., Alvarez, J. A., Cavagnaro, J. B., Guevara, A., Sartor, C., Passera, C. B., Greco, S. (2011). Ser planta en el desierto: estrategias de uso de agua y resistencia al estrés hídrico en el Monte Central de Argentina. Ecología Austral, 21(1), 029–042. https://ojs. ecologiaaustral.com. ar/index.php/Ecologia_Austral/article/view/1294
Xavier, J. B., Andrade, D. B. D., Souza, D. C. D., Guimarães, G. C., Resende, L. V., & Guimarães, R. M. (2019). Morphological, chemical and physiological characterization of Amaranthus spp. Seeds. Journal of Seed Science, 41(4), 478-487. https://doi.org/10.1590/2317- 1545v41n4226286
Zubillaga, M. F.; Repupilli, J. A.; Boeri, P.; Servera, J. A.; Gallego, J. J.; Piñuel, L. (2024). Nutritional quality of amaranth (Amaranthus) silage in response to forage airing and addition of lactic bacteria. Revista de la Facultad de Ciencias Agrarias. Universidad Nacional de Cuyo. Mendoza. Argentina. 56(1): 117-126. DOI: https://doi.org/10.48162/rev.39.128
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.